The Elongated African theory is an evolutionary hypothesis devised by the late Belgian anthropologist Jean Hiernaux. It was introduced in his 1974 book The People of Africa, a work which seeks to explain the existing physical and genetic diversity in Sub-Saharan Africa through various developmental processes. Touting itself as using a then-new non-racial approach, the narrative places an emphasis on environmental adaptation as one of the primary driving forces behind human biological variation.
Hiernaux suggests therein that the swarthy “Caucasoid” (“Hamitic”) phenotypes which typify the Afro-Asiatic-speaking populations in the Horn of Africa and the Moors in the Sahara evolved through interbreeding between, on the one hand, Arabs or Berbers, and on the other, naturally narrow-featured, hypothetical inhabitants of Sub-Saharan Africa that he dubs “Elongated Africans.” To prove his theory, Hiernaux employs a series of logical fallacies, contradictions and factual inaccuracies, many of which were apparent even at the time of publication. For this reason, his Elongated African hypothesis never really took off. It was instead criticized by some of his own colleagues, eventually abandoned by Hiernaux himself, corrected a few years later by more comprehensive anthropometric, craniometric and serological studies, and debunked altogether through ancient DNA analysis. Although now obsolete, the theory remains an instructive example of how excessive post-colonial guilt can easily lapse into faulty science and guesswork.
Development of the theory

Map showing the spread of the Niger-Congo-speaking ancestors of the Tutsi-Hima Bantus and Hutu Bantus from their suggested Original Bantu Homeland on the Nigeria-Cameroon border in West Africa (Jaja et al. (2011)).
Hiernaux was a physician and anthropologist by training. Beginning in the 1950s, he alongside the historian and archaeologist Emma Maquet carried out some of the first excavations in present-day Rwanda, Burundi and the Democratic Republic of the Congo in eastern Central Africa. These Great Lakes territories were at the time under Belgian colonial rule.

Phylogeny of the Niger-Congo language family. Kinyarwanda, the mother tongue of the Tutsi Bantus and their brethren the Hutu Bantus, belongs to the Narrow Bantu branch of the Niger-Congo phylum (Bostoen (2004)).
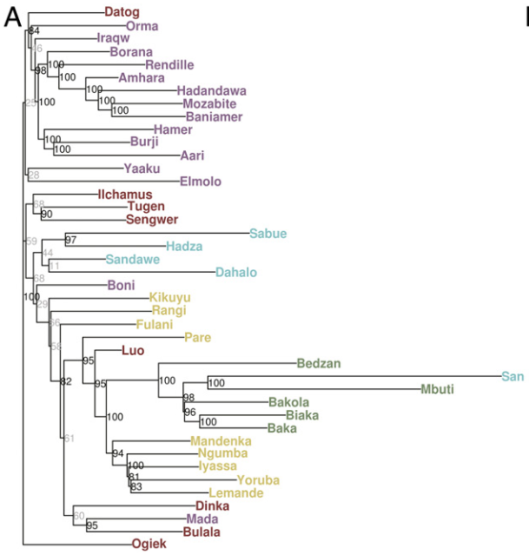
While stationed in the area, Hiernaux also conducted a series of anthropometric studies on the local Tutsi pastoralist, Hutu agriculturalist and Twa hunter-gatherer populations. Although all three groups speak the same Kinyarwanda language (a member of the Bantu branch of the Niger-Congo family), he argued that they each had distinct origins and suggested that these differing ancestral backgrounds could be discerned anthropologically. To this end, in several of his earlier works, Hiernaux insists that a “Hamitic” influence is evident among the Tutsi despite their linguistic, cultural and physical commonalities with the “Bantu” Hutus and “Pygmy” Twa.
This assertion was in part drawn from the Tutsis’ own longstanding oral traditions, as documented by the explorer John Hanning Speke, among others. These local accounts held that the founders of the various Great Lakes kingdoms were “Caucasoid” peoples, who had arrived in the region several centuries prior from either the Horn or North Africa. The Hamitic migrants are then said to have ruled over the local “Negroid” inhabitants, gradually adopted the latter’s Bantu languages, and eventually amalgamated with the autochthones through intermarriage. These stories are embodied in the person of Kintu, a figurative representation of the early Cushitic settlers in the lacustrine region. A. B. Fisher (1904), writing on the territory north of Buganda, states:
The oldest inhabitants of the country related to me how a white man and woman many years ago landed on the right bank of Lake Albert, and settled in Bunyoro. On being questioned as to whether they referred to Sir Samuel and Lady Baker, they replied that they remembered them well, and described them to me. But, said they, these other white people arrived a long, long time ago, and founded their first dynasty. The story is that this man and his wife came down from the Nile, and caused a great sensation, the people all flocking round them, exclaiming, “Kintu ke?” (What is it?). So they named the man Kintu.
As explained in part two of this essay, there is some truth to the above narrative, for there exists today a minor Cushitic paternal influence among the Tutsi Bantus.

Portrait of Kintu and his wife Nambi. Note the couple’s “Caucasoid” appearance. According to A.J. Mounteney Jephson (1893), this is because Kintu and his bride are the “personification of the influential immigrants from Galla countries” in Northeast Africa, who moved south and settled in the Great Lakes region.
-

-
An Oromo man with the cad (“olive-skinned”) phenotype, which is found among the Cushitic peoples. In the colonial period, such individuals were referred to as the “white Galla.” Traditional accounts of the early Cushitic settlers of the Great Lakes region alternately describe the latter as having been “red” or “white” in complexion. Thus, it is likely that at least some of these ancient pastoralists had an cad physiognomy, as many Cushitic peoples still do today.
-

-
An Oromo woman, with an example of the cad or “white Galla” phenotype among Cushitic women.
Hiernaux later distanced himself from his initial writings, apparently in response to intensified tensions between the Tutsis and Hutus. In his 1968 work, while ironically decrying what he termed “classificatory mania”, he still maintained that the Tutsi were biologically distinct from the other Bantu-speaking Great Lakes aborigines, the Hutu and Twa. It was around this period when Hiernaux developed what he would eventually call his “Elongated African” hypothesis.
In the first iteration of his theory, Hiernaux argued that the Tutsis were largely of “Ethiopid” origin (traditionally a synonym for “Eastern Hamite”). This ancestral stock, he wrote, was neither Europoid nor Negroid nor a mixture thereof, but rather a race unto itself. By 1972, Hiernaux would assert that there was no significant exotic component in the Tutsi, suggesting instead that they evolved their physique through genetic adaptation to hot and dry conditions. He would also propose that the Afro-Asiatic-speaking populations of the Horn and the Moors of the Sahara were ancestrally formed through a mixture of Arabs and a mysterious, environmentally-molded “African” stock similar to the Tutsi. Hiernaux thus essentially changed the direction of the gene flow, arguing now that it was the Afro-Asiatic speakers that experienced a Tutsi-like influence rather than vice versa.
-

-
An Algerian Berber man, representing a Maghrebi individual of typical western “Hamitic” type (Keane (1899)).
-

-
A Bisharin Beja man, representing a Cushitic individual of “pure” eastern “Hamitic” type (Keane (1899)).
This second incarnation of his theory was primarily inspired by the work of Hubert Jules Deschamps, a French historian and colonial administrator. Based on anthropometric means and blood work, Deschamps had asserted in 1970 that Somalis, Ethiopians, Moors and certain other groups on the southern rim of North Africa all appeared to have considerable exotic affinities (“Arab” in his nomenclature). He suggested that this ancestral affiliation manifested itself most conspicuously through traits that were not found in combination elsewhere below the Sahara, such as non-kinky hair texture, keen facial features, and oftentimes lighter skin pigmentation. Deschamps, however, conceded that it was impossible to quantify this ancestry without knowing first just what exactly was the nature of the African element in these populations. He also spoke of a Moors-Somali Warsingali “constellation”, and noted that these Afro-Asiatic-speaking groups lived in similar “biotopes” or habitats.

Phylogeny of human stocks in Africa. The Ethiopian “Hamites” and “Mediterranean” peoples of the Horn of Africa and North Africa descend from a common “Proto-Caucasoid” stock (Cole (1954)).
A few years later, the pioneering anthropologist and archaeologist Sonia Cole would invite Hiernaux to write for her Peoples of the World book series published by Charles Scribner’s Sons. Cole had released The Prehistory of East Africa in 1954, a seminal work that provides strong evidence for an early Caucasoid presence in the region as well as in North Africa. She had most recently authored the British Museum of Natural History guide Races of Man (1963), which draws on her many years of field work in Africa and again emphasizes an ancient Caucasoid presence on the continent.

Distribution map of the peoples of Africa, including the “Caucasoid” Hamites and Semites. The Tutsi are among the Bantu-speaking Group of “Negroid” peoples (Cole (1963)).
Against this backdrop, Hiernaux, then serving as the Director of Research at the National Centre for Scientific Research in Paris, would pen his The People of Africa in 1974.
Ancient skeletons in East Africa
Hiernaux starts off by arguing that an ancestral “Elongated African” population can be found in the makers of the Upper Kenya Capsian (Eburran), a Mesolithic hunter-gatherer culture centered in East Africa. He is aware of the Upper Kenya Capsian people’s proposed Caucasoid metrical affinities, measurements which he does not dispute. He also acknowledges that that lithic industry has ties with the coeval Capsian culture of North Africa, whose makers he indicates were gracile “Mediterraneans”:
The makers of the Capsian are less well known physically than the makers of the Ibero-Maurusian. They are Mediterraneans, whose lighter build contrasts with the robustness of the Mechta people. Probably the Berbers are their descendants, with a possible admixture of the Mechta element in some places.
The oldest remains of Homo sapiens sapiens found in East Africa were associated with an industry having similarities with the Capsian. It has been called the Upper Kenya Capsian, although its derivation from the North African Capsian is far from certain. At Gamble’s Cave in Kenya, five human skeletons were associated with a late phase of the industry, Upper Kenya Capsian C, which contains pottery. A similar association is presumed for a skeleton found at Olduvai, which resembles those from Gamble’s Cave[…] The skeletons are of very tall people. They had long, narrow heads, and relatively long, narrow faces. The nose was medium width; and prognathism, when present, was restricted to the alveolar, or tooth-bearing, region. Many authors regard these people as physically akin to the Mediterraneans, hence the label of ‘Caucasoids’ (or European-like) generally attached to them.
Where Hiernaux differs is in the actual origin of those Caucasoid osteological affinities. He suggests that those traits developed in situ in East Africa, independently from the Capsian Mediterraneans in North Africa. The reason why Hiernaux asserts this is because he believes that all of these features can be found in the modern Tutsi Bantus and Maasai Nilotes of the Great Lakes, two populations that he indicates (wrongly, as it turns out) have little-to-no exotic biological influences.
Hiernaux’s first error is in assuming that the Gamble’s Cave skeletal remains of his day were in the same condition as when Louis Leakey first excavated and described them in 1928/29. These fossils were from the start fragmentary, and only two of the five could later be reconstructed. The specimens incurred further damage when the Royal College of Surgeons, where they had subsequently been stored, was bombed during WWII. For this reason, the anthropologist G. Philip Rightmire — who, unlike Hiernaux, had the opportunity to study the remains in the 1970s — correctly notes that no firm conclusions on the fossils’ affinities could by then be ascertained:
Deposits belonging to this “Gamble’s Cave Shoreline” complex have now been dated to between 8000 and 10,000 B.P. Of the five Gamble’s Cave skeletons, only two could be reconstructed, and this job was carried out in England after the material had been sent there from East Africa. Results were certainly far from perfect, owing to warping and crushing of the original bone, and further insult was to follow. The Royal College of Surgeons in London and the skeletal collections housed there received heavy bomb damage during World War II. So by the time that the skulls were transferred to the British Museum (Natural History) in 1948, they were scarcely in mint condition. Skull number 4 is the less well preserved of the two, and all of the base as well as a substantial portion of the facial skeleton are present only in plaster. Distortion renders this specimen quite unfit for measurement. Number 5 also lacks much of the skull base, and the missing parts have been heavily reconstructed. Although these skulls have been called non-Negro in morphology, the evidence is certainly far from clear cut, and any such diagnosis is questionable by virtue of the state of the material alone.

The Cro-Magnon (left) and Gamble’s Cave Number 4 (right) skulls. Note especially the distorted eye orbit shape of the latter, heavily reconstructed cranium (Oschinsky (1963)).
Furthermore, Hiernaux mistakenly assumes continuity in the skeletal record from the makers of “the Upper Kenya Capsian of Gamble’s Cave, Naivasha and Olduvai, who may date to about 4,000 BC; [to] the makers of the ensuing mesolithic Elmenteitan culture of Bromhead’s Site; [to] the remains associated with the neolithic stone bowl culture at Hyrax Hill and Njoro River Cave (dated by carbon 14 to 960 BC), and with the more recent stone bowl culture at Willey’s Kopje, Makalia and Nakuru, which almost certainly date from the Iron Age.”
As we saw, the actual affinities of the fragmentary Upper Kenya Capsian remains are uncertain. Because the Gamble’s Cave Number 4 skull was heavily reconstructed, its morphological status is somewhat conjectural. Oschinsky (1963), for instance, remarks that the “Gambles Cave II, No. 4 skull shows lateral compression which has distorted the shape of orbits, the left zygomatic arch is twice as long as the right one, the basic occipital region is displaced to the left, the cranio-facial juncture has been crushed toward the rear of the neuro-cranium on the right side causing the short right zygomatic arch. The same pressure from the front of the skull has caused the alveolar region to be flattened and the palate to be deepened.”
According to the anthropologist Steven H. Ambrose, the industries at the Njoro River Cave, Makalia and several other Rift Valley sites that Hiernaux indicates were part of the Stone Bowl Culture were also actually later expressions of the Elmenteitan culture:
Extraction of the central incisors may prove significant for confirming correlations between modern and prehistoric cultures. Although this is not exclusively a Nilotic practice, it is most common among modern Nilotic populations in Tanzania and Sudan (Kilma, 1970: 8; Murdock, 1959: 173). The central incisors were removed from all 79 crania recovered from Njoro River Cave, an Elmenteitan cremation burial cave (Leakey and Leakey, 1950: 76). They may therefore have been of Nilotic origin. This practice is also evident in an early Iron Age context at Wiley’s Kopje and the Makalia Burial Site, both located on the western side of the Rift Valley (L. S. B. Leakey, 1935: 95, 107-108). The latter site may represent an Iron Age expression of the Elmenteitan Industry (Chittick et al., in press).

I. M. Lewis excavating a cairn at Gaan Libah in northern Somalia (Lewis (1957)).
The Stone Bowl Culture, also known as the Savanna Pastoral Neolithic, is generally associated with early Southern Cushitic settlers. However, just who exactly were the makers of the Elmenteitan culture is a question that, until recently, was disputed. Louis Leakey and Mary Leakey first analyzed human remains at the Elmenteitan-associated Njoro River site in Kenya. They noted that these specimens had a “non-negro” morphology and were markedly different from the present-day Bantu and Nilotic inhabitants, including the groups with some Cushitic admixture. The Leakeys observed that the Njoro River skeletons had a hyper-leptorrhine (very narrow) nasal index of 47.88. By comparison, Leakey and Leakey (1950)’s lowest scoring Bantu/Nilotic sample, the Tanganyika natives, had a platyrrhine (broad) nasal index of 55.4. Hiernaux (1960)’s Iron Age Rwanda sample was also platyrrhine, with a nasal index of 58.6.

Reconstructed necklaces from the Njoro River Cave, an Elmenteitan site of the Pastoral Neolithic. Similar beads have been found in predynastic Egyptian burials (Cole (1954)).
In 1957, I. M. Lewis excavated a series of cairns at Gaan Libah in northern Somalia, which were radiocarbon-dated to a maximum of 250 years before present. The skeletons buried within the graves were subjected to an anthropometric analysis. In stark contrast to the Bantu/Nilotic samples, the Gaan Libah specimens turned out to be hyper-leptorrhine, with a virtually identical nasal index as the Njoro River individuals (viz. 47.8). This only further validated the Leakeys’ theory on the identity of the original Elmenteitan culture bearers. (Note: the nasal indices above were calculated based on the skulls of the deceased; for the nasal indices of living individuals, see below under Anthropometry). Likewise, after examining 16 beads that were recovered from the Njoro cave, H. Beck opined that “some of them show great resemblance to the pre-dynastic Egyptian work” (Leakey and Leakey (1950)).
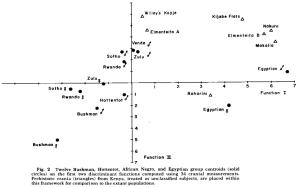
Analysis of human crania from ancient burial sites in the Rift Valley and those of various Bantu, Khoisan and Egyptian groups. The ancient Cushitic Makalia and Baharini samples cluster with the male and female Egyptian samples. The Rwanda male and female samples cluster instead with the Sotho Bantus and other Niger-Congo-speaking populations (Rightmire (1975)).
In 1975, Rightmire compared human crania from ancient burial sites in the Rift Valley with those of various Bantu (including Rwanda), Khoisan (Bushman, Hottentot), and Egyptian (Egyptian E/Gizeh) groups. He observed that several of the analyzed Rift Valley skulls shared greatest affinities with the Egyptian samples; namely, the Baharini, Makalia I, and (albeit tentatively) Elmenteitan F1 specimens. As Rightmire put it, “there is little doubt about Baharini as Egyptian female.” The rest of the skeletons either clustered with the “negro” crania or were in the process of being assimilated into these Bantu and Khoisan groupings. This reflects the gradual replacement of the early Egyptian-related Cushitic settlers in the Great Lakes area with the incoming Bantu/Nilotic populations; the latter peoples now constitute the region’s predominant inhabitants.
Based on these skeletal affinities as well as population dynamics and historiolinguistics, Ambrose correlates the Caucasoid skulls with the first Cushitic pastoralists and the Negroid crania with those of the ancestral Nilotic populations:
Craniometric studies undertaken by Rightmire (1975b, this volume) suggest that among the crania from Neolithic burial sites in the Rift Valley were representatives of Nilotic Negroid populations, as well as those whose closest correlates are found among prehistoric Egyptian populations. Rightmire suggests that many of the Rift Valley crania may represent speakers of a Nilotic language (Rightmire, 1974; 1975a, b). One cannot at this point correlate each individual cranium with an individual industry, due to small sample sizes and lack of precise information on artifactual associations. However, we may be dealing with as many as three distinct physical types: a Nilotic Negroid, a Cushitic “Caucasoid,” and an indigenous Negroid hunting population (Gramly and Rightmire, 1973). Cushites and Nilotes have a history of several thousand years of contact along a major geographic and ecological boundary that follows the Ethiopia/Sudan and northern Kenya/Uganda border regions (Ehret, 1974b). Archaeological and linguistic evidence demonstrates that the region of contact continued south along the Rift Valley, Kenya, to Lake Eyasi, Tanzania. The Eyasi region is a cul-de-sac where representatives of the four major language phyla of Africa — Khoisan (Hadza and Sandawe), Afro-Asiatic (Southern Cushitic), Nilo-Saharan (Eastern and Southern Nilotic), and Niger-Congo (Bantu) — are spoken today. Racial differences have undoubtedly been minimized by intermarriage during the long period of contact. Thus, although there is a strong skeletal evidence for two different Neolithic populations, with Sudanic and Ethiopian origins, respectively, it may never be possible unerringly to correlate skeletal types with industries.
With regard to the affinities of the makers of the Kenya Capsian/Eburran culture, the historian and linguist Christopher Ehret indicates that the “Mediterranean Caucasoid” skulls that have often been associated with this industry likewise appear to have actually belonged to the Cushitic makers of the Savanna Pastoral Neolithic/Stone Bowl Culture. He suggests that the makers of the Eburran culture were instead likely of Khoisan ancestral stock:
The poorly understood Eburran Industry has misled other people on the basis of a spurious, misinterpreted association of “Mediterranean Caucasoid” skeletons (Leakey 1931; Protsch 1975, 1978), which are more likely to be of Highland Savanna PN origin. This association has led some people to postulate an early center of Cushitic speech in central Kenya (Fleming 1964, 1969, 1976: 265; Murdock 1959: 197). Since there are no skeletons that can actually be attributed to pre-Neolithic industries, it is more likely that the makers of this industry originated from a local population with great antiquity. Given evidence for the formerly widespread distribution of Khoisan languages throughout East Africa, it is more likely to have been spoken by Eburran hunter-gatherers and other pre-Neolithic East African populations than an Afroasiatic language.
Correlation of the Savanna PN “Stone Bowl Culture” with Southern Cushitic groups has been the orthodox interpretation for many years (Sutton 1966, 1971, 1973; Odner 1972; Phillipson 1977a). Southern Cushitic speakers are theorized to have been the earliest food producers in East Africa, possessing cattle, sheep, and goats and probably cultivating grain. They would correlate with the Lowland Savanna PN, the first pastoralists, who made stone bowls and Nderit Ware in the Turkana region between 5,000 to 3,000 b.p. and later, at 3,300 b.p., spread to the highlands of Kenya and northern Tanzania.
This interpretation is not controverted by the skeletal evidence amassed by Rightmire (1975), which shows that many of the Neolithic peoples of the Rift Valley have their closest affinities with Egyptian populations. The skeletal evidence, however, also demonstrates the presence of peoples whose closest affinities were with modern Negroid populations and who were not “Mediterranean Caucasoids,” as Leakey proposed (1935). Thus two groups of people, of different racial and geographic origins, were present in the Rift Valley.
This is in keeping with Ambrose’s assertion that there was a third early physical type present in the region alongside the Cushitic Caucasoids and Nilotic Negroids. Leakey (1936) likewise notes “Bushman affinities” for the Later Stone Age (40,000 ybp-2,000 ybp) inhabitants of the Lake Victoria area. In contrast, he emphasizes that the Gumban/Savanna Pastoral Neolithic makers were of “a physical type which is almost European”. The anthropologist Gunter Bräuer reaches a similar conclusion in his large study on the morphological differentiation of anatomically modern humans in Africa:
although the East African highlands probably cannot be regarded as the centre of differentiation of modern man in Africa, this part of the continent does represent an important region which was inhabited by Europid, Negrid and Khoisanid populations in prehistoric times

Distribution of ancestral populations in Africa circa 9,000 BCE (Bräuer (1978)).
Ambrose further confirms the above when he observes that the chronological date proposed by R. Protsch for the cairn burials at Gamble’s Cave is grossly inaccurate. To this end, Ambrose notes that conventional charcoal dates for the older (and thus deeper) Phase 3 layer at the site range from 8,000 to 8,500 years before present. Protsch, however, had mistakenly suggested that the cairns — which were buried in a deposit above the Eburran’s final/most recent Phase 5A layer; Phase 5A was, in turn, situated around four meters above the Phase 3 layer — dated to a similar 8,020 ybp, give or take a few years. Thus, the cairns are in fact chronologically more recent than even the last Eburran cultural phase, and by extension, so are the skeletons within them. The specimens’ “Mediterranean Caucasoid” morphology therefore indeed likely does not represent the general physical type of the makers of the Eburran culture. It instead appears to have arrived in the region later with the Cushitic makers of the Savanna Pastoral Neolithic/Stone Bowl Culture, as Ehret had correctly deduced. Ambrose writes:
Dates sites comprising Phase 5A of the Eburran listed in Table ie show that this phase began before 2900 B.P. and ended well after 2000 B.P.[…] The same reservations apply to Protsch’s date of 8020±260 (Protsch, 1978: 103) for the cairn burial overlying the Eburran 5A horizon at Gamble’s Cave, as this date is inconsistent with conventional charcoal dates ranging from 8500 to 8000 B.P. on Phase 3 in this site from 4 meters below this horizon. The Gamble’s Cave burials actually overlay the Phase 5A horizon (L. S. B. Leakey, 1931: 117), and are thus later than and unrelated to this horizon, and may not represent the Eburran physical type[…] Therefore, conventional dating evidence indicates that the Mediterranean Caucasoid physical type belongs to the Neolithic era.
J. E. G. Sutton makes the same observation, remarking that Leakey himself indicates in his excavation notes that the burial cairns containing the “Mediterranean Caucasoid” skeletons belong to a separate, much later population than the makers of the Kenya Capsian/Eburran:
The ‘Kenya Capsian’ is a blade-and-burin industry, which in North Africa or Europe would be classified as ‘Advanced Palaeolithic’. In fact, Leakey originally called it ‘Kenya Aurignacian’ by comparing it with French materials, later allowing its redesignation as ‘Kenya Capsian’ on account of Maghrebian comparisons. The type-site (indeed the only site with a collection of reasonable size, satisfactorily stratified and at least cursorily described in print) is Gamble’s Cave (the lowest ‘occupation level’), which Leakey excavated in the late 1920s.[…] The base of these deposits has now been radiocarbon-dated (following a test cutting by Glynn Isaac and Ron Clark in 1964) to the seventh millennium B.C. It was here that Leakey had collected, beside the stone tools and waste in quantity, both fish-bones and a broken harpoon, though this later find was not recognized till thirty years after the excavation![…]
Repeatedly in the literature the makers of the ‘Kenya Capsian’ are described as a ‘tall Caucasoid’ or ‘Afro-Mediterranean’ people, a deduction based on examination of burials which Leakey found while digging Gamble’s Cave. Whether this racial attribution is roughly correct or not is irrelevant here. For, as is plain in Leakey’s ‘diagrammatic section’ and notes of his excavation, these burials were placed in a layer well above that containing the true ‘Kenya Capsian’ materials with the fish-bones, harpoon and ‘dotted wavy-line’ potsherd. The skeletons probably belong to a different population several thousand years later. There is therefore no direct evidence of the physical type of the makers of the ‘Kenya Capsian’.
In the early 1980s, the anthropologist and archaeologist Daniel Stiles set out to assess the affinities of ancient human fossils buried in several Savanna Pastoral Neolithic/Stone Bowl Culture sites as well as in Azanian cairns in the Chalbi Desert. Stiles had helped establish the Department of Archaeology at the University of Nairobi during the 1970s, so he was well qualified for the task. He later published three papers on the excavations, Stone Cairn Burials at Kokurmatakore, Northern Kenya (1981), The Azanian Civilization and the Megalithic Cushites Revisited (1984), and The Azanian Civilisation Revisted (2004).

Daniel Stiles unearthing the 3,500 year old (~1,500 BCE) Savanna Pastoral Neolithic mound cairn, which he indicates is “the oldest dated stone structure in East Africa”.
Many of the skeletons that Stiles excavated were complete and in good enough condition for him to measure their standing height. They were generally quite tall individuals of Caucasoid physical type, consistent with oral and written tradition. Stiles was also able to accurately date the cairns that the specimens were interred in. He found three different types of cairns, each belonging to separate epochs. The oldest cairns were of mound type; they dated to around 3,000 years ago, and were evidently made by early Southern Cushites of the Stone Bowl Culture. The next oldest cairns were of platform type; they dated to about 1,000 years ago and thus were probably made by early Eastern Cushites. The third group of cairns were of ring type and dated to around 600 years ago. Unlike the more ancient specimens in the mound and platform cairns, the skeletons within the ring cairns showed some evidence of Nilotic cultural influence since all of them had their lower incisors removed. Stiles therefore suggests that these more recent cairns probably contained the Cushitic ancestors of the Rendille, who are known to have intermarried and exchanged customs over the years with the adjacent Samburu Nilotes, including the practice of incisor extraction.
On one of the Savanna Pastoral Neolithic-associated mound cairns that Stiles excavated, he writes:
The ~3,500 year old (~1,500 B.C.) mound cairn was almost certainly made by Southern Cushitic speaking ‘Stone Bowl’ pastoralists. A stone bowl fragment was found buried at the base of the cairn, and obsidian tools and goat bones were found inside the cairn. The person buried inside measured 190cm tall (about 6’4”), an incredible height for someone of that antiquity.
-
-
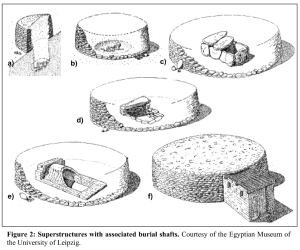
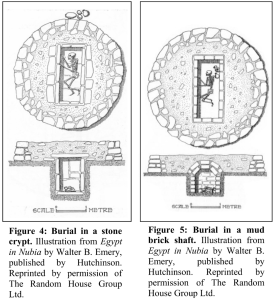
Ancient morturary structures of the C-Group people of Lower Nubia (present-day southern Egypt). These burials consist of a stone crypt (either built above ground or subterranean) or mud-brick vault in which the body of the dead is placed, encircled by a stone ring superstructure made of dry-wall (Hafsaas (2005)).
-
-
Aerial view of typical C-Group burials, encased in a circular stone superstructure. On the left, the deceased rests in a stone crypt, and on the right, the body has been placed in a mud-brick vault (Hafsaas (2005)).
-

-
A ring cairn built by the Cushites of the Pastoral Neolithic at Kokurmatakore, Kenya. This burial, consisting of a grave pit in the center surrounded by a ring of stones, is evidently a remnant of the same kind of funerary structures which the ancient C-Group pastoralists of the Nile Valley would erect (Stiles and Munro-Hay (1981)).
-

-
An enclosed platform monument at Salaxley Valley near Garowe in northeastern Somalia, one of many such ancient burials in the area. Chittick (1975) also reported the existence of similar mortuary structures on the coast at Ras Hafun. Like the ring cairns further south at Kokurmatakore in the Great Lakes region, these ring and platform cairns of the Horn appear to be vestiges of the more elaborate crypt/vault and superstructure burials of the C-Group peoples and Neolithic pastoralists in the Nile Valley. During the subsequent Christian and Islamic eras, masons probably bit-by-bit disassembled the dry-stone walls and roofs, which once encased these graves. This would have given these builders the raw material they needed to construct new mosques, churches and other religious edifices. A comparable process is known to have occurred in medieval Egypt, when the Islamic dynasties repurposed the ancient pyramids’ original gleaming limestone surface in order to erect their grand mosques.
In short, Hiernaux was completely mistaken about the affinities of the various ancient skeletons in East Africa. He erroneously assumed that the Kenya Capsian/Eburran, Elmenteitan and Savanna Pastoral Neolithic/Stone Bowl cultures were all made by a single hypothetical population, his “Elongated Africans”. In reality, the Kenya Capsian/Eburran was likely the work of early hunter-gatherer peoples, who, based on their lithic industry, probably had some cultural (if not demic) contacts with the Capsians of North Africa. Ancient DNA analysis has, moreover, proven that the original makers of both the Savanna Pastoral Neolithic and Elmenteitan cultures were early Cushitic pastoralists (see below under Exotic influences). Additionally, archaeogenetics has revealed that the ancient Nilotes were instead responsible for the Pastoral Iron Age sites. The “Mediterranean Caucasoid” morphology is also associated with only one of those groups, the Cushitic settlers of the Pastoral Neolithic.

Three-dimensional stereogram comparing various modern populations with some early specimens from the Great Lakes, Southern Africa and the Nile Valley (Bräuer (1980a)).
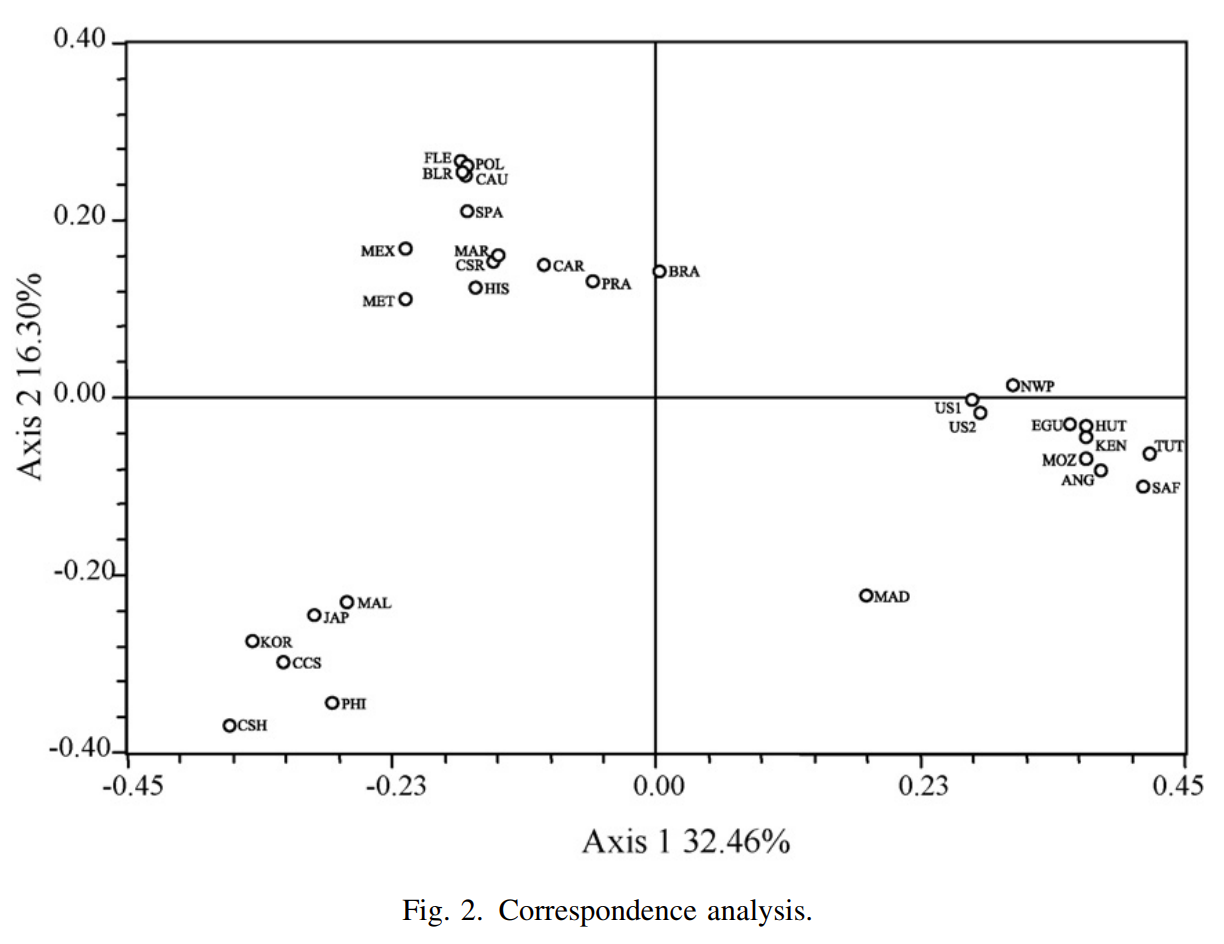
That said, just how do these ancient Caucasoid specimens of the Great Lakes relate to the present-day Afro-Asiatic-speaking populations to the north, in the Horn region? Are they, as Hiernaux postulates, ancestral to Cushitic speakers in general? That is, are they the immediate forebears of the Eastern Cushites (like the Somalis, Afar, Saho, Sidamo and Galla/Oromo), Northern Cushites (Beja), Central Cushites (Agaw), and the largely extinct Southern Cushites? Or are they instead early Southern Cushites alone, as Stiles asserts? Hiernaux does not offer a comparative analysis, but thankfully Bräuer does. Bräuer (1980a) finds that his modern Galla-Somali lumped sample is more closely related to ancient Afro-Asiatic groups from the Nile Valley, particularly the predynastic Egyptians of Naqada and the C-Group peoples of Lower Nubia/Northern Sudan. This is true in all three principal coordinate dimensions; notably, in the first axis, which contains most of the variation between the examined populations (26.2%).
Modern haplogroup and lactase persistence allele analyses provide a similar indication, for they establish that the early Afro-Asiatic-speaking presence in the Great Lakes region was mainly represented by the Southern Cushites (see below under physiognomy and exotic influences). These ancient Caucasoids south of the Horn would eventually be absorbed by the aboriginal hunter-gatherer populations and the incoming Nilotic and Bantu groups.
In part two of the Elongated African fallacy, we demonstrate how we know for a fact that the Caucasoid morphology did not develop in situ in East Africa as Hiernaux had proposed. We also show how the physical anthropology and ecology underpinning his theory are likewise flawed, contradictory and generally inaccurate.
In part one of the Elongated African fallacy, we saw how Jean Hiernaux was initially one of the prime exponents of Hamitic scholarship in African historiography. Under post-colonial duress, he gradually distanced himself from his earlier writings and drew inspiration from the work of Hubert Jules Deschamps. We also saw in detail how and why Hiernaux was mistaken about the affinities of the various ancient human fossils in East Africa, specimens which he erroneously assumed were all of the same physical type. In this second and final part of the Elongated African fallacy, we further demonstrate the inherent flaws and contradictions in his hypothesis by deconstructing the actual physical, serological and environmental evidence that he invokes to support it.
Exotic influences
In his book The People of Africa, Hiernaux includes a brief chapter on biological, cultural and linguistic classifications, where he asserts that his “Elongated African” populations are “much too diverse to form a taxon”. He then devotes an entire chapter to his Elongated African theory, where he also discusses the Nilotes. Hiernaux starts off by defining what exactly he believes physically constitutes an “Elongated African”, as well as which four populations today best represent this putative morphology in its unaltered state:
As already discussed in Chapters 5 and 6, a number of African populations have an elongated body build, with narrow head, face and nose. Their skin is dark (in varying degree), their hair is spiralled, and they have thick but not everted lips. In many of these people, such as the Tutsi of Rwanda and Burundi and related Hima of Uganda, the Masai of the East African steppes and the Ful communities of the Western Sudanic savanna, there is no evidence of an exotic (Arabic or North African) element in their gene pool. Their physical features can best be explained in terms of genetic adaptation to dry heat. Apparently they represent the result of a peculiar evolution in the semi-arid crescent which caps sub-Saharan Africa to the north and north-east.

A Nilotic Maasai man. The Maasai are sometimes termed “Hamiticized Negroes” due to the fact that their Nilote ancestors in the Great Lakes region assimilated some earlier Southern Cushites.
Right off the bat, Hiernaux makes a fundamental factual error that all but invalidates his theory. He mistakenly assumes that the Tutsi-Hima Bantus, Maasai Nilotes and Ful West Africans — his “pure” Elongated Africans — do not have any extraneous physical influences that could account for their more Caucasoid-leaning morphology than other Bantus, Nilotes and West Africans, respectively. In reality, all of these populations have low-to-moderate levels of Afro-Asiatic admixture, which their ancestors acquired through interbreeding with early Cushitic and Berber groups. For this reason, such populations were often referred to in the anthropological literature as “Hamiticized Negroes”, or, if the Hamitic influence was believed to be a bit more salient, as “Negro-Hamites” or “Half-Hamites”. Sonia Cole in her masterwork The Prehistory of East Africa describes the situation thusly:
The Upper Kenya Capsian people were very tall, over 5 feet 10 inches, and had long, narrow skulls with prominent chins and noses. They are apparently of Caucasoid or Mediterranean type, and may be termed ‘Proto-Hamites’ to emphasize their resemblance to the present inhabitants of North Africa and the Horn (the term ‘Hamite’, often used to describe North and North-East African people with marked racial characteristics, should strictly speaking apply to a language group only). Possibly a mixing of this proto-Hamitic type with incoming Negroes (of which there is no sign in Africa during the Upper Paleolithic) gave rise to some of the modern semi-Hamitic negroid peoples, such as the Masai.

Hima Bantus (sitting) and Bairu Bantus (standing). Like their Tutsi Bantu relatives further west in the Great Lakes region, the Hima of Uganda share a close physical and genetic resemblance with the Bantu peoples dwelling near them, the Bairu.

Tutsi Bantu men styling their natural afro-textured hair in the traditional amasunzu coiffure.
Hiernaux, on some level, appears to be aware of this since he mentions various Afro-Asiatic-speaking populations on the continent as “possible sources of ‘Hamiticization’.” He also remarks that the Maasai and Tutsi “have some characteristically Cushitic food habits.” Additionally, in reference to the linguist Joseph Greenberg and the anthropologist George P. Murdock, Hiernaux notes that “the pastoral Masai[…] their language belongs to the Eastern Sudanic class in Greenberg’s classification. However, both their language and culture show a strong Cushitic influence, which makes Murdock describe them as ‘Kushitized Nilotes’.”
-

-
A Maasai warrior. Notice the man’s body-wrap, armband, earrings, trinkets, necklace and general mode of dress, which, according to Oric Bates, is very similar to the traditional attire of the ancient Libyans. We now know that this is because the Maasai and other Great Lakes Nilotes borrowed much of their material culture from the Cushites of the Pastoral Neolithic. The ancient Cushites were closely related to the eastern Libyans, with whom they also shared many cultural aspects. This material culture eventually influenced neighboring Nilotic, Bantu and hunter-gatherer populations they came into contact with.
-

-
An ancient Egyptian slab depicting an eastern Libyan man. Note the figure’s body-wrap, necklace and armband, which the modern Libyans no longer wear. These sartorial elements have instead been taken up by the Maasai, Nandi and other Nilotic peoples inhabiting the Great Lakes region. Consequently, this style of dressing is often misattributed as traditional Nilotic attire, when in fact it is borrowed ancient Libyan/Cushitic culture.
DNA
In the case of the Tutsi and Maasai, a notable Cushitic male influence is indeed evident in their uniparental lineages. Trombetta et al. (2015) observed that around 78% of Tutsis in Burundi and 76% of Maasai in Kenya carry typical Bantu and Nilotic clades. However, a significant minority are M293 bearers (Tutsi=22%; Maasai=24%). M293 is a subclade of E1b1b (E3b), a paternal haplogroup that is most common among Afro-Asiatic speakers in the Horn and North Africa. The M293 sublineage is specifically associated with early Southern Cushites, for it peaks among remnant Southern Cushitic speakers in the Great Lakes and its modern geographical distribution also closely mirrors the historical distribution of the Southern Cushitic languages. More importantly, the earliest occurrences of M293 have been identified in ancient skeletons belonging to the Pastoral Neolithic, a Cushitic-affiliated cultural complex (see below). Thus, we now have genetic confirmation that the subhaplogroup was indeed introduced to the lacustrine region by the Southern Cushites.
-

-

-
Y-DNA haplogroups among Niger-Congo and Afro-Asiatic-speaking populations. The Niger-Congo-speaking Tutsi and Hutu of Rwanda both primarily belong to the E1b1a or E3a clade, which today is the most common paternal lineage carried by Bantu speakers. By contrast, the South Cushitic-speaking Iraqw/Wairak of Tanzania principally carry the E1b1b or E3b haplogroup, a typical paternal lineage borne by Afro-Asiatic speakers (Luis et al. (2004); Luis et al. (2007)).
Although the South Cushitic-linked E1b1b-M293 clade has been detected at low-to-moderate frequencies among Tutsis and Maasai, more extensive Y-DNA analysis has unveiled that the majority of individuals from these communities actually belong to the E1b1a or E3a haplogroup. E1b1a is today the most common paternal lineage among Bantu speakers. Luis et al. (2004) and Luis et al. (2007) report that almost all Tutsis in Rwanda bear this clade (over 75%). This suggests that the Cushitic admixture existing among the Tutsi Bantus is more pronounced in Burundi than in Rwanda. Furthermore, Wood et al. (2005) note a high frequency (50%) of the typical Afro-Asiatic-affiliated haplogroup E1b1b in a sample of Maasai individuals in Kenya. Trombetta et al. (2015), however, observed a lower percentage (37.8%) of all E1b1b subclades in their Maasai sample from the same territory (cf. Supplementary Table 7). Hirbo (2011) reports an even lower E1b1b or E3b percentage of just 16.7% for his Kenyan Maasai cohort. Likewise, Mwema (2011) found that the most common patrilineal STR markers carried by Maasai in Tanzania (51 individuals, the largest Y-DNA sampling of Maasai) are instead assigned to the E1b1a clade. Hence, the Cushitic admixture present among the Maasai Nilotes appears to be focalized in only select areas in Kenya; the “purer” Maasai population in Tanzania and elsewhere in the Great Lakes region shows a considerably lower Afro-Asiatic influence (as also indicated by autosomal DNA analysis; see below).

Y-DNA haplogroups of the Tutsi Bantus, Hutu Bantus and other Sub-Saharan African populations. Most Tutsi individuals (80%) and Hutu individuals (73%) carry derivatives of the E3a or E1b1a lineage like other Niger-Congo-speaking groups, which mainly consist of the basal E3a* clade and the E3a7 subclade (Hirbo (2011)).
Similarly, Hassan et al. 2008 found that around 54% of Ful/Fulbe/Fulani migrants in Sudan are haplogroup R1 carriers. This clade is today the most common paternal haplogroup among males in Europe. The presence in Africa of its R1b-V88 sublineage (which now is mainly borne by Fulani and Chadic speakers) is believed to represent late-glacial period diffusion from the Franco-Cantabrian area of southwestern Europe. This is supported by genome analysis of modern Fulani individuals, who carry a bit of Anatolian Neolithic-related ancestry like that characteristic of Iberian peoples (see Vahaduo Multi analysis here; the Fulani individuals on average bear ~7.6% Anatolian Neolithic admixture, as exemplified by the TUR_Marmara_Barcin_N sample). A further 35% of Ful belong to M78, a subclade of haplogroup E1b1b/E3b that is thought to have originated in Egypt. Hence, contrary to what Hiernaux claims, there is not one but at least two separate sources of exotic influence among the Ful: an ancient Iberian one and a Berber one.
Unlike their Y-DNA/paternal DNA, the mtDNA/maternal DNA of the Tutsi, Maasai and Ful appears to show little exotic influences (cf. Cerný et al. (2006), Castri et al. (2008)). This suggests that the Afro-Asiatic admixture in these populations was instead mainly acquired through the past assimilation of Cushitic and Berber males. In regards to the Tutsi, Göbel et al. (2019) found that almost all individuals in their Rwanda sample belonged to derivatives of the macrohaplogroup L. Much of this mitochondrial variation was also shared with the Kenya cohort. The scientists did, however, detect a minor Cushitic influence in their Rwandan dataset via the presence of the West Eurasian maternal lineages M1a1 (2.6%), N1a1a (1.3%) and K1a (0.6%) as well as the U6a clade:
The Rwandan sample[‘s] […] broader haplogroup pattern, mirrored also by higher MNPD (Table 1), is[…] highly similar to that reported from Kenya.[…] Most of the non-L low-frequency lineages found in the novel datasets are highly informative about human history. Lineages M1(a) and U6(a) are explained as signals of ancient backflow into North Africa from the Mediterranean area in the Early Upper Paleolithic. N1a1a is a low-frequency lineage with a relict distribution likely indicating a Pleistocene dispersal from Arabia. All are reported also in other East and North African populations. The single haplotype of K1a, a lineage found across West Eurasia according to EMPOP, might be attributable to more recent migration to Rwanda and is, intriguingly, shared with the dataset from Somalia.
This stands in sharp distinction to the neighboring Iraqw of Tanzania, a relict South Cushitic-speaking population that has managed to retain significant frequencies of Afro-Asiatic-associated uniparental markers (both Y-DNA and mtDNA) despite being completely surrounded by hunter-gatherer/Bantu/Nilotic communities. In this respect, Hirbo (2011) reports that 55.3% of his Iraqw sample bore the E1b1b paternal clade (of which 51% was assigned to the M293 or E3b6 subclade), with 12.8% carrying the T or K2 lineage. The examined Iraqw individuals also had a moderate incidence of West Eurasian maternal haplogroups (~22%), much of which consisted of the M1a clade (8.7%).

Analysis of HLA antigens among populations in Africa, the Middle East and Europe. Ethiopians, Beja and Sudanese “Arabs” cluster with other Afro-Asiatic-speaking groups inhabiting North Africa and the Middle East, as do Nubians. By contrast, Nilo-Saharan and Niger-Congo-speaking populations cluster together, separately from the Afro-Asiatic speakers (Aamer (2016)).
With regard to HLA antigens, Tang et al. (2000) write:
Cumulative anthropologic and genetic evidence indicates that centuries of intermarriage have largely dissolved the ethnic distinctions originally separating tribes (e.g. Hutu and Tutsi subgroups).
As such, the scientists observed that their Rwandan female sample’s HLA profile was generally unique to it, though broad commonalities could be discerned with certain other populations in Africa. Of these, the Rwandan cohort’s DRB1 and DQB1 haplotypes were again most similar to those of the Kenyan samples, but rather different from those of the West and Central African samples. This finding concurs with the Y-DNA haplogroup data on the Tutsis, for it suggests that they and the Hutus are most closely related to other Bantu populations in the Great Lakes region (e.g. the Kikuyu Bantus, who experienced comparable hunter-gatherer, Nilotic, and Cushitic admixtures as the Tutsi Bantus). Ali et al. (2020) likewise indicate that the Rwanda population’s HLA profile is analogous to those from other territories in eastern central Africa (viz. Kongo Kinshasa, Central African Republic, Uganda). By contrast, Lulli et al. (2009) report on the Fulani/Fulbe that: “In particular, the DRB1*04 allele is absent or rare in all Sub-Saharan African populations, except in the Fulani and in Amhara-Oromo from Ethiopia, where it reaches a frequency close to that of Europeans. These observations are in agreement with the hypothesis that the Fulani’s genetic make-up includes an appreciable Caucasoid component of possible East-African origin, which has been suggested on the basis of their physical features and cultural traditions.” This is also supported by Mohamoud (2006), who notes that “the result of HLA class I and class II antigen frequencies show that the Somali population appear more similar to Arab or Caucasoid than to African populations.”
-
-
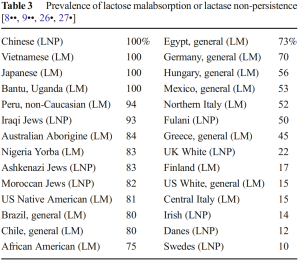
Global prevalence of lactose malabsorption/lactase non-persistence. Many Fulani individuals today are lactose absorbers/lactase persistent, as are most native Europeans. This is a reflection of gene flow from the Franco-Cantabrian area of southwestern Europe, which impacted the ancestral Fulani pastoralist community. Conversely, most of the Egyptian, Jewish, Sub-Saharan African and East Asian samples consist of lactose malabsorbers/lactase non-persistent individuals, due to these populations’ respective agrarian backgrounds (Bayless et al. (2017)).
-
-
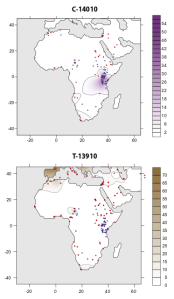
Contour map showing the allele frequency distribution in Africa of the lactase persistence C-14010 allele (associated with Southern Cushites) and T-13910 allele (associated with Europeans). Many Fulani individuals carry the Europe-specific T-13910 variant, which again they acquired through contact between their West African ancestors and earlier European populations (Ranciaro et al. (2014)).

DNA analysis of the long-horned “Ankole” cattle kept by Tutsi-Hima herders indicates that they are a variety of “Sanga,” the most common cattle breed owned today by Niger-Congo and Nilo-Saharan-speaking pastoralists in Africa. Sanga cattle is a crossbreed of the “Hamitic Longhorn” or taurine cattle of ancient Egypt (Bos taurus) and the zebu cattle of the Indian peninsula (Bos indicus). The Hamitic Longhorn, now almost extinct, has long since been replaced by the zebu and the camel among Afro-Asiatic speakers in Northeast Africa and by different varieties of the hardier Sanga cattle among Niger-Congo/Nilo-Saharan speakers elsewhere. The lone exception is among Fulani pastoralists of the Sahel, who keep herds of N’Dama cattle. N’Dama is the only bovine variety in Africa today that still has predominant taurine ancestry, consistent with the Fulani’s suggested links to the ancient Nile Valley (Kwon et al. (2022); Tijjani et al. (2022)).
Further evidence of exotic influence among the Maasai and Ful can be seen in the lactase persistence alleles that they carry. Tishkoff et al. (2007) observed that up to 58% of the Maasai bear the C-14010 variant. According to Breton et al. (2014), this lactose tolerance allele originated among Afro-Asiatic-speaking pastoralists in East Africa (likely Southern Cushites), who then spread it in the surrounding area all the way to the Khoe-inhabited parts of Southern Africa. This postulated diffusion is now supported by ancient DNA analysis (see below), which has identified the C-14010 variant in an early Cushitic specimen associated with the Pastoral Neolithic. This points to a rapid expansion of lactose tolerance in East Africa, just as has been observed in Europe. Similarly, Lokki et al. (2011) found that around 37% of Fulani in Mali carry the T-13910 mutation, the most common lactase persistence allele among Europeans. DNA analysis of the long-horned cattle raised by Fulani pastoralists, moreover, indicates that the “N’Dama” breed they keep is largely the same “Hamitic Longhorn” or taurine variety (Bos taurus) of ancient Egypt (cf. Kwon et al. (2022); Tijjani et al. (2022)). Altogether, this is a reflection of the aforementioned gene flow from the Franco-Cantabrian area of southwestern Europe, which impacted the ancestral Ful.
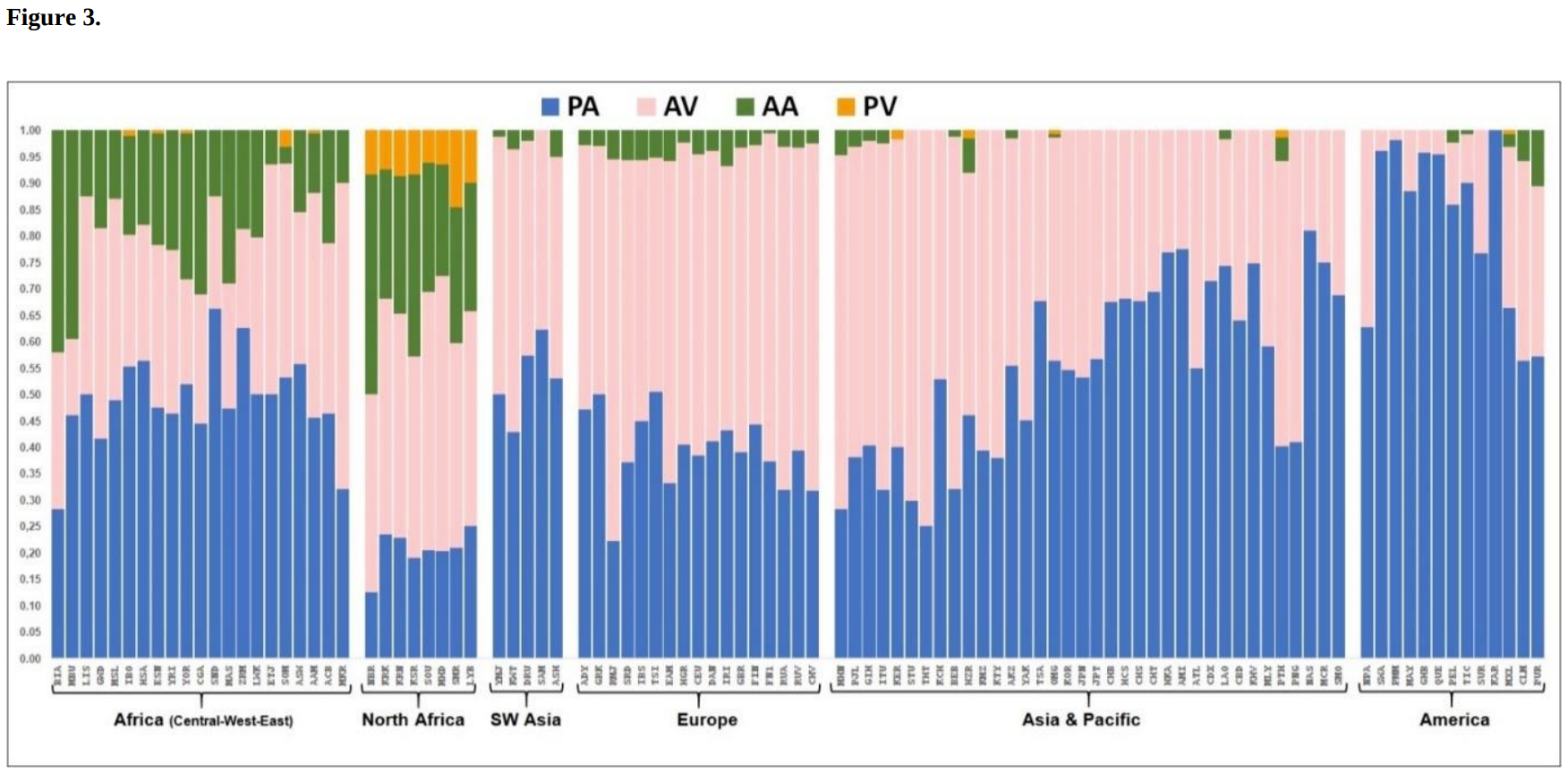
Worldwide allele frequencies of the TAS2R38 gene, which is associated with bitter taste perception. Most of the Somali and Ethiopian Jew individuals as well as the other Afro-Asiatic-speaking samples from the Middle East carry the global PA variant, followed by the Europe-centered AV allele. By contrast, most of the Maasai Nilotes bear the global PA variant, followed by the Africa-centered AA allele (Mourali-Chebil et al. (2022)).
As regards bitter taste perception, which is genetically inherited, analytical data indicates that Maasai individuals share similar allelic variants as other Nilo-Saharan and Niger-Congo speakers. Mourali-Chebil et al. (2022) observed that most of the Maasai Nilotes they examined bore the global PA variant of the TAS2R38 gene, followed by the Africa-specific AA allele. By contrast, after the generic PA variant, most of the Somalis, Ethiopian Jews and other Afro-Asiatic-speaking samples from the Middle East were found to carry the Europe-centered AV allele, with few individuals bearing the AA variant.

Global frequency distribution of the various isoforms of the APOE gene. After the globally-distributed APOE3 allele (ε3), most Tutsi Bantus and other native populations of central Africa bear the Africa-centered APOE4 or ε4 variant (nearly 40%). In contrast, Afro-Asiatic speakers from the Horn of Africa and Arabian peninsula that have been examined almost all carry the APOE3 isoform, which climaxes among Amerindian groups as well as Basques and Sardinians in southern Europe (Abondio (2021)).
Concerning variability in the apolipoprotein E (APOE) gene, which influences longevity in humans, Abondio (2021) reports that, after the APOE3 isoform (ε3) which prevails in all contemporary populations, her Tutsi Bantu cohort, Aka Pygmies and other native samples from central Africa carried among the highest worldwide frequencies (close to 40%) of the Africa-centered APOE4 isoform (ε4). By contrast, the Afro-Asiatic-speaking individuals from Ethiopia and the Arabian peninsula whom she studied almost exclusively bore the APOE3 variant (>85%), which peaks among Sardinians and Basques of southern Europe as well as among Amerindian populations. Abondio indicates:
ε3 is currently the most frequent isoform in all modern populations, peaking at 94% in the Hutterite people of Canada, 90% in Mayas, 88% in the Basque and Sardinian populations, and 86% in Han Chinese people. Isoform ε4 shows the maximum percentage in a range of indigenous populations of Central Africa (40% in Aka Pygmies, 38% in Tutsis), Oceania (49% in the Hui population of New Guinea) and Central America (27% for the Huychol people of Mexico). Moreover, a decreasing North-South gradient (Figure 3.5.1 and Appendix A1) characterizes this isoform in Europe (5% to 10% in the Mediterranean basin; 16% in Central and Western Europe; 23% in the Scandinavian peninsula, with peaks of 31% in Finland) and China (Zekraoui et al. 1997; Corbo and Scacchi, 1999; Singh et al. 2006; Hu et al. 2011; Ojeda-Granados et al. 2017). Isoform ε2 has an inconsistent distribution, with peaks in Southeast Asia, Australia, Africa (19%) but is absent Native American groups (Singh et al. 2006).

Density map of sickle cell in Africa and the Middle East (Ridley (2003)). Most modern Niger-Congo-speaking populations carry sickle cell alleles at variable frequencies, including the Tutsi Bantus and their Hutu brethren in the Great Lakes region (estimated at 1%-5% and 5%-15%, respectively; cf. Bain (2006)). Although they today inhabit the malarial zone, the Afro-Asiatic-speaking populations in Northeast Africa generally do not bear any sickle cell variants. Sickle cell mutations are, however, present among some individuals in Northwest Africa and the Middle East, primarily in the form of the Arab-Indian haplotype (cf. Al-Ali et al. (2020)).
With respect to sickle cell anemia, Bain (2006) reports that 1%-5% of Rwandan Tutsi, 5%-15% of Rwandan Hutu, and 1.5%-26% of Burundi inhabitants have the associated Haemoglobin S (HbS). This trait is generally not found among the Cushitic-speaking populations of the Horn of Africa (Bain notes a ~0% HbS frequency in Djibouti and Somalia), but it does occur among a significant minority of modern Bantus and other Niger-Congo-speaking groups. Prendergast et al. (2018) further indicate that their early Bantu sample from the Deloraine Farm in Kenya’s Rift Valley (dated to c. 1170-970 years before present) lacks any sickle cell alleles (cf. Table S12). This suggests that the Tutsi Bantus and Hutu Bantus did not derive their sickle cell variants directly from the Iron Age Bantus, but instead from the same, more recent source as other contemporary Bantu speakers in the Great Lakes region.
-

-
Analysis of GM immunoglobulin allotypes carried by various populations in Africa. The Cushitic-speaking (Issa Somalis of Djibouti) and Ethiosemitic-speaking (Amhara and Tigray of Ethiopia) samples from the Horn of Africa have affinities with the Berber and Arabic-speaking populations from North Africa, due to shared European GM haplotypes (Coudray et al. (2004)).
-

-
Analysis of GM/KM immunoglobulin allotypes carried by various populations in Sub-Saharan Africa. The Tutsi, Hutu and Twa samples from Rwanda/Burundi primarily cluster with other Niger-Congo-speaking groups, as “the Tutsi and Bantu show close genetic associations” (Weber et al. (2000)).
In terms of the GM/KM immunoglobulin allotype system, Weber et al. (2000) analysed Tutsi, Hutu and Twa individuals from Rwanda/Burundi and found that “the Tutsi and Bantu show close genetic associations.” Coudray et al. (2004) likewise observed that their Fulani sample from Senegal clustered with other Niger-Congo-speaking populations. On the other hand, their Cushitic-speaking (Issa Somalis of Djibouti) and Ethiosemitic-speaking (Amhara and Tigray of Ethiopia) samples from the Horn of Africa showed affinities with their Berber and Arabic-speaking samples from North Africa. Coudray et al. suggest that this is because these Afro-Asiatic-speaking populations share European GM haplotypes:
L’axe 1 de cette deuxième AFC permet de mettre en évidence 2 grands groupes de populations. Un groupe comprenant les populations nord et est africaines : Berbères du Maroc, d’Algérie, de Tunisie ; Touaregs d’Algérie et populations est africaines d’Ethiopie et de Djibouti. Cet ensemble se divise en trois sous-groupes selon l’axe 2 : Touaregs d’Algérie, Berbères et Amhara Tigrai et Issas d’Afrique de l’Est. L’autre grand groupe rassemble les populations sub-sahariennes (Mali, Sénégal, Côte d’Ivoire, Nigeria et République Centrafrique). La position de ces groupes aux extrémités de l’axe 1 révèle l’importante distance génétique les séparant. Cette distance se traduit par une différenciation entre les haplotypes Gm « européens » (Gm21,28;1,17;.., Gm21,28;1,2,17;.., Gm5*;3;23 et Gm5*;3;..) pour le groupe nord et est africain et les haplotypes Gm « africains » (Gm5*,28;1,17;.., Gm5*;1,17, Gm5,6,11,24;1,17;.., Gm15;1,17;.., Gm5,6,10,11,14,28;1,17;.. et Gm5,6,10,11,14;1,17;..) pour les populations sub-sahariennes. Nous pouvons aussi remarquer que cette distinction de 2 ensembles de populations africaines se superpose à une distinction géographique de part et d’autre du Sahara. Le désert ne représente pas pour autant une barrière aux gènes car on trouve chez les Berbères nord-africains environ 20 % d’haplotypes « sub-sahariens ».
Axis 1 of this second AFC makes it possible to highlight 2 large groups of populations. A group comprising the North and East African populations: Berbers from Morocco, Algeria, Tunisia; Tuaregs from Algeria and East African populations from Ethiopia and Djibouti. This set is divided into three sub-groups along axis 2: Tuaregs from Algeria, Berbers and Amhara Tigrai and Issas from East Africa. The other large group brings together the sub-Saharan populations (Mali, Senegal, Ivory Coast, Nigeria and the Central African Republic). The position of these groups at the ends of axis 1 reveals the great genetic distance between them. This distance results in a differentiation between the “European” Gm haplotypes (Gm21,28; 1,17; .., Gm21,28; 1,2,17; .., Gm5 *; 3; 23 and Gm5 *; 3 ; ..) for the North and East African group and the “African” Gm haplotypes (Gm5 *, 28; 1.17; .., Gm5 *; 1.17, Gm5,6,11,24; 1.17; .., Gm15; 1,17; .., Gm5,6,10,11,14,28; 1,17; .. and Gm5,6,10,11,14; 1,17; ..) for the sub-Saharan populations. We can also notice that this distinction of 2 sets of African populations is superimposed on a geographical distinction on either side of the Sahara. However, the desert does not represent a barrier to genes because we find among the North African Berbers about 20% of “sub-Saharan” haplotypes.
-

-
Global autosomal STR affinities. Somalis cluster with other Afro-Asiatic-speaking populations in North Africa and the Middle East (Steele et al. (2014)).
-

-
Global autosomal STR affinities. Tutsis (TUT) cluster with Hutus (HUT) and other Bantu-speaking populations in the African Great Lakes (Simms et al. (2010)).
— Global autosomal STR affinities. The Tutsi Bantus again cluster with the Hutu Bantus and other Niger-Congo-speaking individuals. By contrast, the northern Sudanese (SUD) group with Afro-Asiatic-speaking Egyptian (EGY) and Omani (OMN) individuals (From top left, clockwise: Coudray et al. (2006); Simms et al. (2008); Shepard and Herrera (2006b); Shepard and Herrera (2006a)).
In terms of autosomal DNA — which is inherited from both parents (and thus often used to gauge overall ancestry), and studied by analyzing single nucleotide polymorphisms (SNPs) and autosomal short tandem repeats (autosomal STRs or microsatellites) — researchers have again noted a moderate Afro-Asiatic influence among the Maasai and Ful. Autosomal SNP data suggests that this genetic introgression was derived from early Cushitic and Berber peoples, who were absorbed by the Maasai’s Nilotic and the Ful’s West African ancestors, respectively (cf. Dobon et al. (2015); Henn et al. (2012)). Cherni et al. (2016) and Hodgson et al. (2014) estimate that the Maasai of Tanzania and Kenya (Kinyawa), respectively, have on average 12% and 21% of ancient Cushitic admixture, while the Fulani have around 28% of ancient Berber admixture (Table S6).
-

-
Inferred ancestral components of the Fulani, Maasai and other global populations. The Fulani individuals (Afro-Berbers) are predominantly of West African ancestry (Niger-Congo=55%), with significant West Eurasian admixture mostly derived from Berber groups (Maghrebi=28%). Their analogues, the African Qatari individuals (Afro-Arabs), are predominantly of West African ancestry (Niger-Congo=30%) with significant Nilo-Saharan admixture (Nilo-Saharan=20%), as well as West Eurasian admixture primarily derived from Arab groups (Arabian=9%), Caucasus/Iranian groups (Eurasian=9%), and Cushitic groups (Ethio-Somali=7%). The Maasai individuals are predominantly of Nilotic ancestry (Nilo-Saharan=41%) with significant West African admixture mostly derived from Bantu groups (Niger-Congo=16%), as well as West Eurasian admixture primarily derived from Cushitic groups (Ethio-Somali=21%) (Hodgson et al. (2014), Table S6)).
-

-
Structural genome analysis of Afro-Asiatic, Niger-Congo, Nilo-Saharan and Khoisan-speaking populations in Africa. At K=2, the Fulani sample is genetically heterogeneous: some Fulani individuals are similar to the Luo Nilotes and share minor ancestry with the Afro-Asiatic-speaking populations (~15%), whereas other Fulani individuals are similar to the Datog Nilotes and share moderate ancestry with the Afro-Asiatic-speaking groups (~30%) (Scheinfeldt et al. (2019)).
-
-
Population tree estimating Fst genetic distance between ethnic groups in Africa. The Fulani sample clusters with the Kikuyu, Rangi and Pare samples (i.e., Niger-Congo-speaking populations with moderate admixture from Afro-Asiatic-speaking groups). In contrast, the South Cushitic-speaking Iraqw sample clusters with other Afro-Asiatic-speaking populations. This suggests that the Iraqw have retained substantial Cushitic ancestry despite intermingling with adjacent hunter-gatherer/Bantu/Nilotic groups (Scheinfeldt et al. (2019)).

Genome analysis of contemporary Fulani individuals and various ancient and modern global samples. At K=6, the analysed Fulani individuals form two distinct genetic clusters: a FulaniA group that is distinguished by higher non-Sub-Saharan African admixture (consisting of an Iberomaurusian-related orange component, a Pre-Pottery Neolithic-related azure component, and a Balkan European-related red component), and a FulaniB group that mostly harbors Niger-Congo/Nilo-Saharan-related ancestry (yellow component) like the neighboring Wolof, Mande and other Niger-Congo/Nilo-Saharan speakers. The Berber samples appear to represent the source population that contributed most of the non-Sub-Saharan African ancestry borne by the Fulani, as these Afro-Asiatic speakers carry all of the aforementioned ancestral elements. Additionally, the Berber individuals harbor some extra Iran Neolithic-related admixture (dark blue component). They seem to have acquired this genome element after the contact period between their Berber ancestors and the ancestral Fulani because the contemporary Fulani samples do not bear this component. Furthermore, although the Afro-Asiatic speakers from the Horn of Africa share most of the same ancestry as the Berber samples, these Cushitic and Ethiosemitic-speaking individuals are differentiated by minor Late Stone Age (c. 3000-1000 BCE) Great Lakes hunter-gatherer admixture (light blue component); they also do not show any trace of the Balkan European red element. Overall, this supports the traditional interpretation that most of the Fulani’s non-Sub-Saharan African admixture was derived from a Berber-affiliated source population(s) (D’Atanasio et al. (2023)).

Genome analysis on the Vahaduo Admixture JS program’s Single function. Fulanis from Ziniare and peninsular Arab bedouins can be modeled as source populations for Mozabite Berbers and other Maghrebi individuals. The ancestral composition of the Mozabite Berbers here consists of around 75% Fulani-derived ancestry and 25% peninsular Arab Bedouin-derived ancestry. However, this estimate is inaccurate since the modern Fulani Ziniare are not a “pure” West African source population. The average Fulani Ziniare individual bears about 25% Eurasian ancestry, which was specifically acquired through admixture with Maghrebis (similar to the percentage of Cushitic-derived Eurasian admixture, which is now carried by the Maasai Nilotes in eastern Africa). What this Vahaduo Single analysis does establish is that, compared to other West African populations, there is considerable gene sharing between the Fulani Ziniare and Maghrebis. This is consistent with Maghrebi gene flow into the ancestral Fulani population.

Genome analysis of Fulani individuals from Cameroon and Sudan, as well as other modern samples from Africa (Bird et al. (2023)). Both examined Fulani groups almost exclusively derive their Eurasian ancestry from a Morocco Berber-related source (dark blue component). They completely lack the signal of ancient Chadic ancestry, which consists of a composite Eurasian ancestry of Cushitic and Tunisian Maghrebi origin (pink and light blue components). By contrast, this genetic signature of ancient Chadic heritage is found among contemporary Chadic speakers in northern Cameroon, among Sahelian groups like the Kanuri and Kotoko, and reaches a frequency peak among the Shuwa/Baggara “Arabs” of Cameroon (who likely originally spoke an Afro-Asiatic language of the Chadic branch). It is interesting to note that, despite inhabiting western Africa, the Shuwa “Arabs” and Cameroon Sahelian samples also bear Nilo-Saharan admixture (purple component). This supports the suggestion that the Shuwa migrated to their present location from the eastern Sahara and Nile Valley, where their Afro-Asiatic-speaking ancestors would have interacted with local Nilo-Saharan speakers before spreading westward. All in all, the study concurs with our Vahaduo genome analysis above, which detected a clear affinity between the Fulani samples from Ziniare, Burkina Faso and Berber individuals. We can now confirm that this association is indeed primarily attributable to ancient Berber gene flow into the Fulani population.
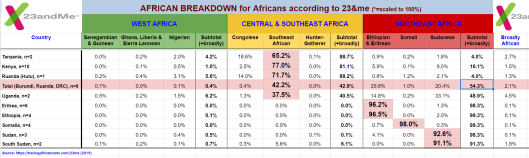
Published autosomal STR analysis on the Tutsi indicates that they are biologically related to the Hutus and other Bantu populations, with little exogenous affiliations (Simms et al. (2008); Simms et al. (2010); Shepard and Herrera (2006a); Shepard and Herrera (2006b); Coudray et al. (2006)). Although no such peer-reviewed autosomal SNP work on the Tutsi-Hima has yet been published, available data on their Hema congeners from the Democratic Republic of the Congo points to a similar Cushitic admixture as found among the Maasai Nilotes (~20%; cf. Xing et al. (2010)). Similarly, autosomal SNP analysis conducted by 23andme, AncestryDNA and other genetic testing companies has traced most of these Tutsi individuals’ ancestry to Niger-Congo/Nilo-Saharan-related forebears (~70% on average); the remainder consists of Cushitic admixture (~30%), which was derived from earlier Afro-Asiatic speakers from Northeast Africa whom the Tutsis’ Bantu/Nilotic ancestors assimilated. These observed predominant Niger-Congo/Nilo-Saharan affinities are in line with a DNA test taken by the Rwandan leader Paul Kagame. According to the Financial Times, although various different admixture strains were detected, the Tutsi politician ultimately shared ancestral links with the neighboring Hutu, consistent with both groups’ spoken Bantu language:
Mr Kagame has taken a DNA test that, he says, reveals him to be of particularly complex genetic mix. The implication, he says, is that he, the ultimate symbol of Tutsi authority, has some Hutu in his genetic make-up.
This is in stark contrast to the Cushitic and Ethiosemitic speakers to the north in the Horn of Africa, who instead share close autosomal SNP (Hodgson et al. (2014), Supplementary Text S1; Dobon et al. (2015); Almarri et al. (2021), Table S4) and autosomal STR ties with other Afro-Asiatic-speaking communities in North Africa and the Middle East (cf. Steele et al. (2014), the largest global autosomal STR analysis).
(*N.B. Using the mAUTO formula, which Stefflova et al. (2009) developed to infer autosomal DNA from known mtDNA and Y-DNA haplogroup frequencies, we may surmise that the Tutsi Bantus of Rwanda bear under 5% total Eurasian ancestry. Since this estimate is around 15 percentage points lower than what is usually projected for them (see estimates below), it would appear that the uniparental lineage frequencies of the Rwandan Tutsi have been shaped by a founder effect. For further details on the mAUTO formula, see Can the Y-DNA and mtDNA of a population be used to infer its autosomal DNA?.)
-
-
Autosomal SNP analysis of Tutsis by the genetic testing company 23andme traces most of these individuals’ ancestry to Niger-Congo/Nilo-Saharan-related forebears (~70% on average). 23andme primarily assigns this ancestry to the Bantu-associated Southeast African cluster (42.2%) and the Nilotic-associated Sudanese cluster (20.4%). The Tutsis’ remaining admixture mainly consists of Cushitic gene flow (~30%), which was derived from earlier Afro-Asiatic speakers from Northeast Africa whom their Bantu/Nilotic ancestors assimilated (Tracing African Roots).
-
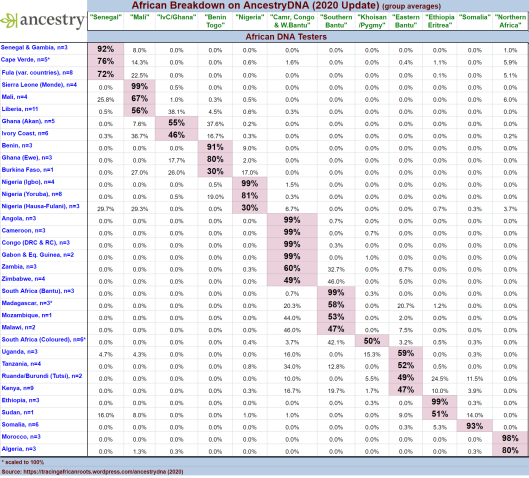
-
Autosomal SNP analysis of Tutsis by the genetic testing company AncestryDNA traces most of these individuals’ ancestry to Niger-Congo/Khoisan/Pygmy-related forebears (~64% on average). As with other Bantu-speaking populations in the Great Lakes region, AncestryDNA primarily assigns this ancestry to the Eastern Bantu cluster (49%), Cameroon, Congo & Western Bantu cluster (10%), and Khoisan/Pygmy cluster (5.5%). The Tutsis’ remaining admixture consists of Cushitic gene flow (~36%), which was derived from earlier Afro-Asiatic speakers from Northeast Africa whom their Niger-Congo/Khoisan/Pygmy ancestors assimilated (Tracing African Roots).
-

-
23andme ancestry test results for a Tutsi Bantu from Rwanda. This individual shares most of his/her ancestry with other Southern East African Bantus (44.8%). He/she also has some admixture from Nilotic peoples from the Sudan area (16.2%). Additionally, this person bears admixture from Cushitic peoples from Ethiopia & Eritrea (34.2%).
-

-
23andme ancestry test results for a Hutu Bantu from Rwanda. Like his/her Tutsi brethren, this individual shares most of his/her ancestry with other Southern East African Bantus (71.7%). However, he/she has comparatively little admixture from Nilotes (2.1%). Additionally, this person bears minute admixture from Cushitic peoples of the Horn region (1.8%).
-
-
Admixture analysis of Tutsi Bantus and other global populations. The examined Tutsi individuals trace most of their ancestry to Sub-Saharan ancestral sources (over 70% on average), comprising Niger-Congo (blue component), Nilo-Saharan (dark green component), Pygmy (pink component), and hunter-gatherer (red component) genome elements. They also have some Eurasian-related admixture derived from Cushitic peoples (~30% of the light green component, which peaks among the Southern Somali sample). This concurs with the commercial genetic tests above of Bantu-speaking Tutsi individuals..
-
-
Global autosomal STR affinities. The Tutsi Bantus (TUT) cluster with the Hutu Bantus (HUT) and other Niger-Congo-speaking individuals. This is consistent with the ancestry results above from 23andme, AncestryDNA and other genetic testing companies, all of which assert Bantu origins for the Tutsi people (Simms et al. (2008)).
-

-
Admixture analysis detecting around 20% of non-African ancestry in the Hema sample. Most of that admixture was derived from earlier Cushitic settlers, whom the Hema’s ancestors assimilated in the Great Lakes region. The Nilo-Saharan-speaking Hema of the Democratic Republic of the Congo are closely related to the Bantu-speaking Hima of Uganda and Tutsis of Burundi and Rwanda, and appear to have once belonged to the same Bantu/Nilotic community. Ergo, it is possible that the Tutsi-Hima have, like their Hema brethren, also acquired minor Cushitic admixture (Xing et al. (2010)).
-

-
Admixture analysis detecting around 30% non-African ancestry in the early Nilotic samples of the Pastoral Iron Age (PIA), excavated in the Rift Valley of Kenya. Most of that West Eurasian ancestry was derived from the Cushites of the Pastoral Neolithic (PN), earlier inhabitants of the Great Lakes region whom the PIA Nilotes absorbed. The ancient Cushitic settlers also introduced the E1b1b and E2 paternal haplogroups, which are now carried by a minority of Nilotes/Bantus/hunter-gatherers in the lacustrine area (Almarri et al. (2020)).
-

-
Admixture analysis detecting around 30% non-African ancestry in the Matangai Turu (c. 750 BP) and Congo Kindoki (c. 150 BP) samples, two early specimens excavated in the Democratic Republic of the Congo. It is tempting to ponder whether such West Eurasian-admixed individuals were ancestral to the local Hema (Wang et al. (2020), Supplementary Material). However, uniparental marker analysis indicates that the only non-Bantu-associated haplogroup which the Congo Kindoki samples carry is the R1b1 lineage, a paternal clade which is not found among the Hema. This suggests that the Hema derive instead their West Eurasian admixture from the Cushitic-admixed Nilotes of the Pastoral Iron Age (Wang et al. (2020), Table S10).
-

-
Y-DNA haplogroups carried by the Hema of the Democratic Republic of the Congo. Many Hema individuals bear the E2-M41 (39%) and E1b1b-M293 (28%) clades. These paternal lineages were first introduced to the Great Lakes region by Cushitic speakers associated with the Pastoral Neolithic cultural complex. This, along with the Hema’s Nilo-Saharan language, supports the view that they directly descend from the Cushitic-admixed Nilotes of the Pastoral Iron Age (Wood et al. (2005), Appendix A).

Genome analysis of Northern Somalis, Oromos, Ethiopian Jews and other global populations, including Maasai. Most of the Northern Somali individuals, who were born in the northeastern Puntland region of Somalia (hailing from the Majerteen Darod clan), cluster in a position parallel to but separate from the North African/Arabian cluster. A few of the examined Northern Somalis are outliers, grouping instead with either the Ethiopian Jew or Oromo samples or shifting toward samples from South Asia. All of the Sub-Saharan African samples are distinct from the Afro-Asiatic-speaking populations from the Horn, North Africa and the Middle East. However, the Maasai Nilotes and Kikuyu Bantus do pull toward the Afro-Asiatic speakers, due to past contact between their Bantu/Nilotic ancestors and neighboring Cushitic groups (Osman and Jonasson (2022)).
In 2018, Prendergast et al. analyzed the DNA of various ancient skeletons in East Africa. The researchers found that the Maasai and other Nilotes in their sample set genetically clustered with the Pastoral Iron Age (PIA) specimens, whereas the Afro-Asiatic-speaking populations grouped with the Pastoral Neolithic samples (PN; comprising the Savanna Pastoral Neolithic and Elmenteitan cultures). They further observed that the Pastoral Iron Age individuals were characterized by ancestry related to modern Nilotic speakers in Sudan, while the earliest Pastoral Neolithic specimens derived most of their ancestry from northeastern Africa/Levant. Additionally, the scientists noted some gene flow from the PN specimens into the later PIA and Maasai samples, consistent with the documented Cushitic influence on the language and culture of the Maasai Nilotes. The researchers therefore suggest that the Pastoral Iron Age individuals were ancestral to the Maasai and other present-day Nilo-Saharan populations in the Great Lakes, and that the Pastoral Neolithic specimens instead represent the forebears of the Afro-Asiatic speakers:
PN individuals, including Elmenteitan and those within the heterogenous SPN category (whom we refer to as “other PN”), mostly form a tight cluster near present-day Afro-Asiatic speakers, with a small number of modest outliers, including the two individuals buried at Prettejohn’s Gully, whose ear-lier date (~4000 BP) coincides with the initial limited spread of herding into the area. Finally, five Iron Age individuals are shifted to the left in the PCA: four PIA individuals toward Nilotic speakers, and an IA child from Deloraine Farm (I8802)—the earliest agricultural site in Kenya’s Rift Valley (32)—toward western Africans and Bantu speakers.[…]
four PIA individuals spanning an ~800-year period show greater affinity to present-day Nilotic speakers and are associated with an influx of Sudan (Dinka)-related ancestry. Similarities between archaeologically and ethnographically documented material culture suggest that PIA sites may be associated with ancestors of present-day Kenyan Nilotic speakers such as the Kalenjin or Maasai (32, 47). Both the PIA individuals and present-day Maasai retain substantial components of PN-related ancestry, showing that the ancestry composition of PIA and more recent pastoralists reflects mixture with previously established herder groups in eastern Africa.
Wang et al. (2020) later reanalysed the ancient Cushitic pastoralists. Thanks to their improved and more broad-based dataset, including several new Pastoral Neolithic samples from Kenya, the researchers were able to more accurately determine the settlers’ ancestral composition. Like Prendergast et al., they formulated a three-way admixture model using Levantine Chalcolithic, Dinka and Mota specimens as hypothetical ancestral populations to the ancient Cushitic herders. However, admixture analysis confirmed instead that the early Cushitic pastoralists had almost identical ancestry proportions as modern Afro-Asiatic-speaking North African groups: they derived around 80% of their genome from a West Eurasian ancestral population, and had just 5%-10% of supposed admixture from Nilo-Saharan peoples. Thus, the ancient Cushitic pastoralists appear to have belonged to a transplanted North African population(s), which gradually moved southwards and settled in East Africa (cf. Supplementary Material).
-

-
Structural genome analysis of the ancient Cushitic pastoralists of East Africa. The earliest of these settlers have been found to derive around 80% of their ancestry from a North African/Middle Eastern ancestral population (red component). At the time of their initial arrival in East Africa, these Cushitic pastoralists presumably would have borne even higher non-African ancestry (~90%) since they would have lacked forager admixture (blue component), which appears to be indigenous to the region (Wang et al. (2020), Supplementary Material).
-

-
Structural genome analysis detecting a shared West Eurasian ancestry at K=5 for the Cushites of the Pastoral Neolithic, the Epipaleolithic Iberomaurusian specimens from Taforalt, early Neolithic individuals from Ifri n’Amr or Moussa, late Neolithic samples from Kelif el-Boroud, and Chalcolithic period specimens from the Levant (Wang et al. (2020), Supplementary Material).
Wang et al. (2022) reported that a Kerma period individual dating from the Middle Kingdom (c. 4000 year old), who was excavated at the Kadruka site in Upper Nubia (northern Sudan), has a genome that “is genetically indistinguishable from that of early Neolithic eastern African pastoralists located 2500 kms away.” The scientists note further that “our findings are consistent with established models for the southward dispersal of Middle Nile Valley pastoral populations to the Rift Valley of eastern Africa, and provide a possible genetic source population for this dispersal.” Hence, it now has been confirmed that the Cushites of the Pastoral Neolithic indeed belonged to a transplanted ancient North African population, and one with a predominant West Eurasian ancestry at that. (For more details, see Genetic affinities of the Cushitic, Ethiosemitic and North Omotic-speaking populations of the Horn of Africa.)
To find out for ourselves what exactly are the autosomal DNA affinities of the Tutsi-Hima and their Hutu kinsmen, we conducted our own genome analysis of Tutsi, Hima and Hutu individuals using the Vahaduo Admixture JS program. Our Target samples mostly consisted of 23andme and AncestryDNA customers, who posted online their converted and scaled Global25 coordinates. For our Source proxies, we used all of the ancient individuals listed on Eurogenes‘ official Global25_PCA_scaled datasheet. Our results showed that, in terms of proximal ancestry (i.e. recent ancestry), the Tutsi and Hima largely descend from the Cushitic-admixed Nilotes of the Pastoral Iron Age or PIA. Prendergast et al. (2018) analysed the remains of PIA specimens and note that these ancient “PIA individuals spanning an ~800-year period show greater affinity to present-day Nilotic speakers and are associated with an influx of Sudan (Dinka)-related ancestry[…] PIA sites may be associated with ancestors of present-day Kenyan Nilotic speakers such as the Kalenjin or Maasai.” Here are the results for a typical Tutsi individual from Rwanda (see here for the full proximal analysis results for all of the Tutsi-Hima and Hutu individuals in the datasheet):
-
-
Vahaduo Single analysis of a Tutsi from Rwanda, estimating proximal or recent ancestry. This Tutsi individual has a predominant Bantu/Nilotic-associated ancestry (nearly 70%), much of which was derived from the Cushitic-admixed Nilotes of the Pastoral Iron Age (KEN_Pastoral_IA). He/she also shows evidence of additional pulses of admixture with the Cushites of the Pastoral Neolithic (TZA_PN, KEN_Pastoral_N KEN_Pastoral_N_Elmenteitan, and KEN_MoloCave_1500BP elements), as well as some extra gene flow from Bantu (COG_NgongoMbata_220BP, ZAF_400BP, UGA_Munsa_500BP and KEN_IA_Deloraine elements), Nilotic (KEN_Kakapel_900BP and KEN_Kakapel_300BP elements), and hunter-gatherer populations (CMR_Shum_Laka and KEN_LSA elements).
-
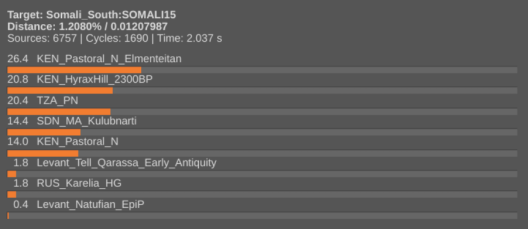
-
Vahaduo Single analysis of a southern Somali, estimating proximal or recent ancestry. This Somali individual has a predominant Cushitic-associated ancestry (~97%), a majority of which was derived from the Cushites of the Pastoral Neolithic (KEN_Pastoral_N_Elmenteitan, KEN_HyraxHill_2300BP, TZA_PN, and KEN_Pastoral_N elements) and a minority of which was derived from the medieval Kulubnarti specimens from Sudan (SDN_MA_Kulubnarti element).
To determine the overall ancestral composition of the Tutsi, Hima and Hutu populations, we then performed another Vahaduo Single analysis, this time using as our Source proxies the “purest” Sub-Saharan African samples (i.e. least Eurasian-admixed) and the “purest” Eurasian samples (i.e. least Sub-Saharan African-admixed) available on the Global25_PCA_scaled datasheet (the methodology followed is described here). Our results showed that, in terms of distal ancestry (i.e. remote ancestry), the Tutsi, Hima and Hutu each traced most of their ancestry to the Kakapel_300BP specimen, which is the “purest” known ancient Nilo-Saharan sample. Here are the results for a typical Tutsi individual from Rwanda (see here for the full distal analysis results for all of the Tutsi-Hima and Hutu individuals in the datasheet):
-
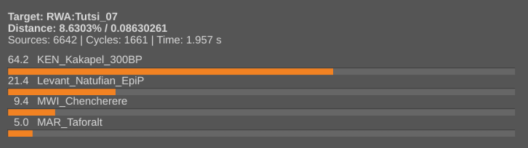
-
Vahaduo Single analysis of a Tutsi from Rwanda, estimating distal or remote ancestry. This Tutsi individual has a predominant Sub-Saharan African ancestry (~75%), a majority of which was derived from the “pure” Nilotes excavated at the Kakapel site in Kenya (KEN_Kakapel_300BP component) and a minority of which came from local hunter-gatherers (MWI_Chencherere component). He/she also bears additional West Eurasian admixture of close to 20% (Levant_Natufian_EpiP component), which was derived from Cushitic peoples whom his/her Nilo-Saharan ancestors assimilated. Additionally, this Tutsi individual has some residual North African Iberomaurusian admixture of around 5% (MAR_Taforalt component).
-
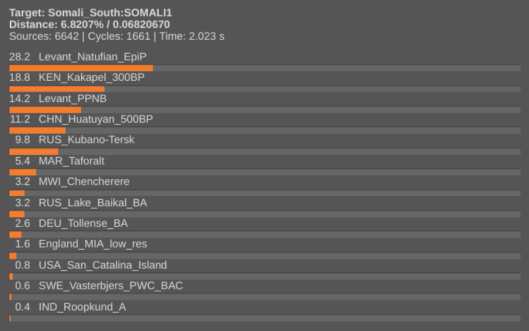
-
Vahaduo Single analysis of a southern Somali, estimating distal or remote ancestry. This Somali individual has a predominant Eurasian ancestry (over 70%), consisting of majority West Eurasian elements (Levantine Natufian-related components: Levant_Natufian_EpiP and Levant_PPNB, as well as European Steppe-related components: RUS_Kubano-Tersk, RUS_Lake_Baikal_BA, DEU_Tollense_BA, England_MIA_low_res and SWE_Vasterbjers_PWC_BAC) and a minority East Eurasian element (East Asian component CHN_Huatuyan_500BP). Additionally, he/she bears some minor Sub-Saharan African admixture (under 25%), which is comprised of a Nilo-Saharan element (KEN_Kakapel_300BP component) and an indigenous hunter-gatherer element (MWI_Chencherere component). This individual also carries a residual North African Iberomaurusian admixture of about 5% (MAR_Taforalt component).
-

-
Vahaduo Single analysis of another Tutsi individual from Rwanda. This analysis includes among its 6000+ comparative Source populations an ancient pastoralist from Skhirat, a site in northern Morocco. Like the Bantu-speaking Tutsi sample above, this Tutsi individual also carries a predominant Sub-Saharan African ancestry (over 75%). Additionally, he/she shows no trace of the Levant-associated Skhirat component, but does carry minor Levantine Natufian ancestry (~18%).
-
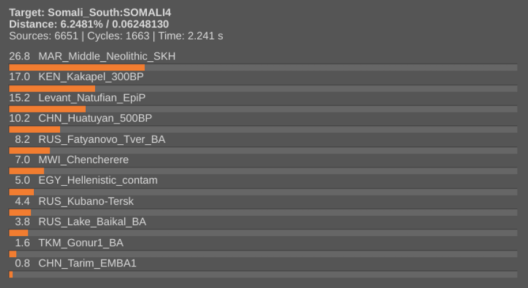
-
Vahaduo Single analysis of another South Somali individual. This Cushitic-speaking person bears a predominant Eurasian ancestry (over 70%), which includes the ancient Levantine component that typifies the ancient pastoralists of Skhirat (~26%). (For more details on these ancient Skhirat pastoralists, see Ancient DNA from Sudan.)
(*N.B. The Vahaduo Single analyses above of the Tutsi, Hima and Hutu individuals should be viewed with discretion since, unlike our Vahaduo Single analyses of the southern Somali individuals (which are based on unscaled or raw Global25 coordinates), these analyses are predicated upon scaled Global25 coordinates. Scaled coordinates are less accurate, filtered versions of unscaled/raw coordinates. As such, they capture less of the genetic variation that an individual carries. We have, nonetheless, made use of these scaled coordinates in our analyses because currently they are the only coordinates of Tutsi-Hima and Hutu individuals that are publicly available. If/when unscaled or raw Global25 coordinates are ever made accessible, we shall update our analysis to reflect this real, unfiltered output.)
The Vahaduo Single analyses above of Tutsi and Hima individuals are also supported by genetic distance analysis. When we carry out a Vahaduo Distance investigation with a focus on proximal or near ancestry, these Tutsi-Hima samples all have a closest genetic distance to the Cushitic-admixed Nilotes of the Pastoral Iron Age. This is in sharp contrast to the South Cushitic-speaking Iraqw, who, despite having sustained considerable hunter-gatherer/Nilo-Saharan/Niger-Congo admixture, are genetically nearest to the Cushites of the Pastoral Neolithic. This confirms our earlier observation that the Iraqw are Cushitic peoples who experienced significant hunter-gatherer/Nilo-Saharan/Niger-Congo admixture, whereas, oppositely, the Tutsi-Hima are Nilo-Saharan/Niger-Congo peoples who experienced some moderate Cushitic admixture.
-

-
Genetic distance analysis of modern Tutsi and Hima Bantus using the Vahaduo Admixture JS program. All of the examined Tutsi-Hima individuals show a clear preference for the Cushitic-admixed Nilotes of the Pastoral Iron Age. This supports the view that the Tutsi-Hima trace most of their proximal or recent ancestry to these earlier Nilotic individuals.
-

-
Genetic distance analysis of modern Cushitic, Ethiosemitic and North Omotic speakers using the Vahaduo Admixture JS program. All of the examined Afro-Asiatic-speaking individuals show a clear preference for the Cushites of the Pastoral Neolithic or the analogous medieval Kulubnarti Nubian specimens. This confirms that these Afro-Asiatic speakers trace most of their proximal or recent ancestry to earlier Cushitic peoples.

Vahaduo Distance analysis of South Somali and Kenyan Somali individuals. All of these Cushitic speakers show closest genetic distance to other Afro-Asiatic-speaking individuals from the Horn vicinity (just 1%-3% genetic differentiation), including samples of other Cushitic (Oromo, Afar, Agaw, Rendille, Iraqw), Ethiosemitic (Amhara, Tigray, Ethiopian Jew), and North Omotic (Wolayta) speakers. Unlike the Iraqw, the Tutsi-Hima do not cluster with the Horn region’s Afro-Asiatic speakers, nor do the Maasai or any other Nilo-Saharan/Niger-Congo/Khoisan populations. This highlights the fact that the Tutsi-Hima are peoples of Bantu/Nilotic/hunter-gatherer ancestral origin who received moderate Cushitic admixture, whereas, oppositely, the Iraqw are people of Cushitic ancestral origin who received Bantu/Nilotic/hunter-gatherer admixture.
In order to better quantify and describe our findings, we ended by running an analysis using the Vahaduo program’s Multi function. Our examined Tutsi and Hima individuals all wound up bearing a predominant Sub-Saharan African ancestry averaging 72%, a majority of which belongs to the “pure” ancient Nilo-Saharan Kakapel_300BP component (~64%) and a minority of which is derived from indigenous East African hunter-gatherers, as represented by the “pure” forager cohort MWI_Chencherere (~8%). The rest of the Tutsi and Hima individuals’ ancestral makeup comprises Eurasian admixture (~22%, almost entirely consisting of Levantine Natufian-associated gene flow) and North African Iberomaurusian/Taforalt admixture (almost 6% on average), with no trace of ancient Niger-Congo-associated admixture (0% of the COG_Kindoki230BP component). By contrast, we detected around 31% of the COG_Kindoki230BP element in our Hutu samples but zero percent of the Taforalt component. This is in marked distinction to the Vahaduo Single proximal analysis above, which did identify a Bantu affiliation of about 10% in most of the examined Tutsi and Hima samples. Since Vahaduo Single analyses capture the exact ancestral makeup of an individual(s) and Vahaduo Multi analyses present that same data in a streamlined and simplified format, we may conclude that persons of Tutsi-Hima background do, in fact, carry some Bantu ancestry, albeit considerably less than what usually has been projected for them.

In brief, genome analysis indicates that the Bantu-speaking Tutsi, Hima and Hutu groups of the Great Lakes region are all closely related peoples. As such, the scholar Mahmood Mamdani (2020)‘s research — which asserts that the dichotomy between the Tutsi and Hutu is primarily one of social caste rather than of different ancestral origins — appears to be correct. On a broader scale, peer-reviewed autosomal STR studies as well as commercial genetic testing link the Tutsi-Hima and Hutu populations with other Niger-Congo-speaking communities inhabiting western Africa. However, our own independent genome analysis has found that, like their Nilo-Saharan-speaking Hema relatives in the Democratic Republic of the Congo, the Tutsi-Hima and Hutu actually descend from a common Nilo-Saharan-affiliated source population. In the archaeogenetic record, this ancestral community is best represented by the 300 BP sample from the Kakapel site in Kenya. The main difference between the Tutsi-Hima and the Hutu seems to be that the Tutsi-Hima had some later contact(s) with pastoralists from northern Africa (as evidenced by the presence among Tutsi-Hima individuals of both the Levantine Natufian and North African Iberomaurusian/Taforalt components) and received comparatively less gene flow from Bantu populations, whereas the Hutu had more intensified interactions with Niger-Congo-speaking groups (as evidenced by the greater frequency of the COG_Kindoki230BP component among Hutu individuals).
Our genome analysis above also seems to support the Kenyan historian Bethwell Ogot (1974)‘s assertion, in regards to the Tutsi-Hima, Maasai and other Bantu/Nilotic peoples that have some Cushitic admixture, that:
the lighter-skinned inhabitants of northern and north-eastern Africa should properly be classified ‘Caucasoid’, or, where necessary, ‘semi-Caucasoid’ or ‘Negroid-Caucasoid crosses’. But such descriptions would not extend to East African pastoralists such as Maasai, Karamojong, Turkana, Hima and Tusi[…] These peoples are decidedly Negroid, though some of them may have incorporated a small admixture of Caucasoid blood in their ancestry.
Physiognomy
With regard to skin pigmentation, Hiernaux contradicts himself again. He affirms above that the Tutsi are dark and specifies elsewhere in his book that they are “very dark skinned”. At one point, he goes as far as to suggest that:
In skin colour, the Tutsi are darker than the Hutu, in the reverse direction to that leading to the Caucasoids.
Yet, in his earlier Hamitic-centered work, Hiernaux maintains that the Tutsi have a lighter complexion than the Hutu. He further asserts that those inhabiting Rwanda are darker-skinned than their brethren in Burundi. This is quite telling since, per Luis et al. (2004), the “darker” Tutsi in Rwanda appear to have a significantly lower Cushitic male influence than Hiernaux’s “lighter” Tutsi in Burundi (they instead are almost all haplogroup E1b1a/E3a carriers, like the average Hutu and most other Bantus). As it is very unlikely that the Tutsi and Hutu incurred a diametric change in coloration within the span of a few years, it follows that Hiernaux’s sudden about-face was motivated by something other than the scientific data.
Global distribution of the derived Ala111Thr/rs1426654 allele, which is associated with lighter skin pigmentation. This SLC24A5 gene variant today has frequencies of around 60% or greater among the Afro-Asiatic-speaking populations in the Horn and North Africa.
Hiernaux makes another error here when he associates a priori dark skin pigmentation with non-Caucasoid ancestry. Ancient DNA analysis has, in fact, confirmed the opposite. That is, most Caucasoid populations — like all early humans — were until recently dark brown-skinned because the alleles linked with lighter coloration are only a few thousand years old. Various pre-Neolithic specimens from Europe with otherwise unexceptional genomes, including ancient individuals from Spain and Greece, were thus found to still have ancestral variants in several skin pigmentation genes. Consequently, even if the early Afro-Asiatic-speaking peoples that the forebears of the Tutsi Bantus, Maasai Nilotes and Ful West Africans encountered had been swarthy, that alone would not necessarily mean that they were not Caucasoids.
As it turns out, these ancient Afro-Asiatic speakers probably instead had light brown complexions. We know this from ancient DNA analysis of early Cushitic settlers in East Africa. Two Pastoral Neolithic individuals (a 2300 year old sample from Hyrax Hill and a 1500 year old sample from Molo Cave, both in Kenya) were tested for the SLC24A5 gene’s derived allele, Ala111Thr or rs1426654. This variant is associated with lighter skin pigmentation, and both specimens wound up carrying it (modern Europeans have this main allele and several additional minor ones; East Asians have their own separate variant). These ancient Cushitic herders also passed on the derived allele to their descendants in the Horn of Africa since the Afro-Asiatic speakers in that region today carry it at high frequencies (cf. Wang et al. (2020), Table S7). Furthermore, ancient DNA analysis of Cushitic individuals of the Pastoral Neolithic cultural complex has found that they bore the derived allele at the APBA2 locus (2 out of 2 examined specimens or 100%), which likewise confers lighter skin pigmentation. Some individuals also carried the derived rs1800404 variant at the OCA2/HERC2 locus, an allele associated with lighter eye color (2 out of 2 examined Pastoral Neolithic specimens from the Lukenya Hill site). These alleles are today still common among Cushitic-speaking groups. By contrast, the early Nilotes of the Pastoral Iron Age were found to bear the ancestral alleles at the SLC24A5, APBA2 and OCA2/HERC2 loci, which are associated with darker skin pigmentation and eye color (Wang et al. (2020), Table S7). (For further details, see What complexion were the ancient Cushitic settlers of eastern Africa? and I have read that some Cushitic peoples have blue eyes. Was this trait inherited from the ancient Cushites?.)
(*N.B. Lazaridis et al. (2022) conducted a comprehensive analysis of phenotypic traits borne by ancient individuals excavated in Europe and Asia. The scientists report that most of their Chalcolithic period sample from Israel (85.7%) had intermediate skin pigmentation, similar to their ancient Minoan cohort from Bronze Age Crete (80%) (cf. Supplementary Materials). According to Wang et al. (2020), this Chalcolithic Levant sample is the best-fitting proxy for the distal source of the West Eurasian ancestry which defines the Cushites of the Pastoral Neolithic. This suggests that these early Cushitic settlers indeed would have been relatively light-complected at the time of their initial arrival in eastern Africa.)

A Somali man (Hawiye clan), with the “Caucasoid” or European-like physiognomy typical of “pure” Cushitic peoples. Because of the close osteological affinities between skeletons belonging to the Pastoral Neolithic culture of eastern Africa and those of modern Afro-Asiatic speakers in the Horn region, anthropologists surmised that these peoples shared ancestral roots. This conjectured link has been confirmed by genomic analysis since contemporary Cushitic, Ethiosemitic and North Omotic-speaking individuals trace most of their proximal/recent ancestry to the Cushitic makers of the Pastoral Neolithic (see genetic analysis above).
In terms of hair form, Hiernaux is more consistent. He writes that “outside of the serological field, hair form appears to be the characteristic which shows the most sharply marked contrast between sub-Saharan Africa as a whole and surrounding areas[…] over most of the subcontinent, spiralled hair is the only category to be observed”. On this basis, he notes that the Tutsi and Maasai have spiralled/kinky hair.

Somali artist Hani UK (Isaaq clan), representing an individual of “pure” Cushitic stock. Note the marked difference in physiognomy between her and the “pure” Tutsi Ange Kagame (shown below), particularly in terms of hair form. This disparity is due to the fact that the Tutsi-Hima are ultimately of Bantu/Nilotic origin, with low-to-moderate Cushitic admixture.
-

-
Gobweyn woman (Cushitic-admixed Bantu). The Gobweyn (Gobauìn) are descendants of the earliest Bantu arrivals in the Horn. This long-term contact with Cushitic peoples is reflected in their language and culture, and oftentimes in their physiognomy (including hair texture) as compared to their “purer” Bantu counterparts.
-

-
Rosalie Gicanda, the last Tutsi queen of Rwanda (moderately Cushitic-admixed Bantu). The Tutsi of the Great Lakes region have moderate amounts of admixture from earlier Southern Cushites, whom their Bantu forebears assimilated. This Cushitic influence is reflected in their tall stature and anthropometric traits, as well as in certain borrowed cultural elements. However, the Tutsi have retained the original Niger-Congo language, skin pigmentation and hair texture of their Bantu ancestors.
-

-
Mushunguli woman (Bantu). The Mushunguli (Musciungullo) are descendants of slaves that were brought to the Horn from southeastern Africa during the 18th century. They represent today the majority of Bantu peoples in Somalia. Most still speak their native Niger-Congo languages, and are generally unmixed with their Cushitic neighbors. Consequently, Mushunguli individuals have also largely retained their original Bantu physiognomy.
By contrast, Hiernaux is well aware that this is not the characteristic hair form of the Afro-Asiatic-speaking populations in the Horn; especially the northern Cushitic groups like the Somali, Afar and Beja:
At the opposite end of the scale, the lowest frequencies of spiralled hair in sub-Saharan Africa have been observed in Ethiopia and Somalia, with a minimum in the Somali.
-

-
A Tutsi Bantu man with the amasunzu hairstyle. Hair morphology analysis indicates that all Tutsis and Hutus have kinky hair texture.
-

-
A Hutu Bantu man with the amasunzu hairstyle.
-

-
A Fulani man. Unlike the Tutsi Bantus and Maasai Nilotes, the Fulani (Fulbe) of the Sahel sometimes have soft-textured frizzy or curly hair. This is due to the fact that the Fulani experienced greater exogenous gene flow, in this case from both ancient Berber and Iberian populations.
-

-
A Tutsi Bantu man with a variation of the Amasunzu hairstyle. It has been conjectured by some writers that this coiffure was meant to emulate the Khepresh or Blue Crown of ancient Egypt.
-

-
Statue of the ancient Egyptian Pharaoh Ramesses II, shown wearing the Khepresh crown.
-

-
Mummy of the ancient Egyptian Pharaoh Ramesses II, with his actual soft-textured, straight red hair visible.
-

-
A Hima Bantu woman. Like their Tutsi Bantu kinfolk, the Hima also exclusively have kinky hair texture.
-

-
A Tutsi Bantu woman with another variation of the amasunzu hairstyle.
-

-
Sculpture of the ancient Egyptian queen Nefertiti (18th Dynasty). Some commentators have opined that the statue’s head form was meant to represent the afro-textured hair of the Tutsi Bantus and other “Negro” peoples.
-

-
Ange Kagame, daughter of the Tutsi President of Rwanda Paul Kagame. Tutsi-Hima women also often attach braid extensions to their natural afro-textured hair.
-
-
Mummy of the ancient Egyptian noblewoman “The Younger Lady,” mother of the 18th Dynasty Pharaoh Tutankhamun. The Egyptologists Marianne Luban and Joanne Fletcher have identified this monarch as Nefertiti. Note her elongated skull caused by artificial cranial deformation, which some members of the ancient Egyptian Amarna royal family used to practice.
-

-
Reconstruction of the mummy of The Younger Lady (left) and statue of Nefertiti. The resemblance between the two figures is striking, consistent with the association of The Younger Lady with Nefertiti. Thus, the unusual head shape of the Nefertiti sculpture above is meant to represent the Younger Lady’s confirmed artificial cranial deformation rather than kinky hair like that of the Tutsi-Hima Bantus.
-

-
President of Rwanda Paul Kagame (Tutsi). Among the Tutsi-Hima, individuals with a more Cushitic-influenced physiognomy can occasionally be found; more often in their ruling class than among commoners. However, this foreign influence is limited to anthropometric traits and never includes attributes such as non-kinky hair texture. This suggests that the Tutsi-Hima’s ancestors were Bantus who absorbed some Cushitic peoples rather than vice versa.
-

-
Brian Kagame, son of Paul Kagame. Brian’s physiognomy shows fewer exotic influences than his father. This is more typical for the Tutsi-Hima Bantus, as they have less Cushitic admixture than the Maasai Nilotes (~20% on average).
-

-
President of Uganda Yoweri Museveni (Hima). The Hima Bantus on average have a bit more foreign admixture than do the Tutsi Bantus. However, ultimately, these two sibling populations are physically and genetically most closely related to other Bantu peoples inhabiting the Great Lakes region.
-

-
Former President of Burundi Michel Micombero (Tutsi). Like many Tutsi-Hima individuals with less extraneous admixture, Micombero has largely retained the Bantu physiognomy of his Niger-Congo-speaking ancestors.
-

-
The singer Stromae, born to Tutsi Bantu and European parents. In physiognomy, he is intermediate between Bantu and European peoples, with a slightly greater pull toward the latter. This is due to the fact that his Tutsi parent already bears some Eurasian admixture (~20%), which was derived from earlier Cushitic peoples whom the Tutsi-Hima’s Bantu ancestors absorbed. Stromae would therefore have inherited around 10% of that foreign admixture, resulting in a more “Caucasoid”-leaning physiognomy than is typical among mixed Bantu-European individuals.
-

-
Tutsi Mwami (King) Kigeli V Ndahindurwa (center) with fellow Tutsi Bantus. Morphological analysis of the skeletal remains of an earlier Rwandan king, Cyirima Rujugira (r. 1675 – 1708), reports that he shared ties with both the Tutsi and Hutu. This is consistent with our genetic analysis (discussed above), which has found that the Tutsi and Hutu ultimately have common ancestral origins, with both groups descending from the same Nilo-Saharan source population.
-

-
Former President of Rwanda Juvénal Habyarimana (Hutu), closely resembling Tutsi-Hima individuals of “purer” Bantu ancestral stock.
-

-
African-American actor Bill Duke. Like many African-Americans/Afro-Caribbeans, Duke bears a close resemblance to Tutsi individuals. This shared likeness underlines the fact that the physiognomy of the Tutsi-Hima Bantus was formed through a similar intermixture between a Niger-Congo/Nilo-Saharan (“Negro”) ancestral base and some minor non-African foreign elements. In the case of the Tutsi-Hima, this intrusive Eurasian gene flow was introduced to their Bantu-speaking ancestors by Cushitic outsiders.
-

-
Tutsi Bantu women au naturel.
-

-
Tutsi beauty contestants in the Miss Rwanda pageant, representing the most attractive women in their Bantu community. The hair modifications (weaves, wigs, relaxing) and skin lightening regimens that typically precede such contests and other modeling engagements serve to give the Tutsi participants a more exotic appearance, approaching that of the Berber-admixed Fulani in Ziniare, Burkina Faso.

African-American singer Jaguar Wright, closely resembling average Tutsi-Hima Bantu women in terms of mesorrhine facial features, dark skin pigmentation, attenuated prognathism, and natural afro-textured hair. This likeness again underlines the fact that the Tutsi-Hima Bantus are of Niger-Congo/Nilo-Saharan origin like African Americans (~75%), with both groups also sharing a similar proportion of ancillary Eurasian admixture (~25%).

A Shuwa (Baggara) “Arab” woman. Genome analysis indicates that the Shuwa/Baggara of the Sahel carry a composite Eurasian ancestry, which consists of components of Cushitic and Berber origin. This composite Eurasian signature appears to be the signal of ancient Chadic ancestry since it is also borne at lower frequencies by Chadic speakers (cf. Bird et al. (2023)). As such, the Shuwa/Baggara seem to have originally spoken an Afro-Asiatic language of the Chadic branch. This explains why the Shuwa/Baggara are physically, genetically and culturally much closer to Afro-Asiatic speakers than are Sub-Saharan tribes like the Tutsi-Hima, despite the fact that the Tutsi-Hima’s Bantu ancestors received low-to-moderate gene flow from Cushitic peoples.
-

-
Tutsi and Hutu Bantu children.
-

-
Nilotic Dinka men from South Sudan. Genome analysis indicates that Tutsi-Hima individuals trace most of their proximal or recent ancestry to the Cushitic-admixed Nilotes of the Pastoral Iron Age. As such, the Tutsi-Hima share ancestral roots with the Dinka and other Nilotic peoples.
— Hutu and Tutsi Bantu individuals from Rwanda. To the layperson, the average Hutu and Tutsi are virtually indistinguishable, owing to their shared Bantu origins. They are, in fact, officially classified as the same Bantu ethnic group, known as Banyarwanda in Rwanda and Banyarundi in Burundi. However, some minor physical differences do exist between the two, mainly in terms of height and nasal index.
— Peoples of Nilotic origin. Top row, from left to right: Dinka man, Nuer man, Tutsi man, Maasai man. Bottom row, from left to right: Dinka woman, Nuer woman, Tutsi woman, Maasai woman. Genome analysis indicates that the Tutsi-Hima and the Maasai of the Great Lakes region trace most of their proximal or recent ancestry to the Nilotes of the Pastoral Iron Age. This shared Nilo-Saharan ancestry readily explains why the Tutsi-Hima, despite today speaking a Bantu language, most closely resemble the Maasai and other Nilotic peoples.
Because Hiernaux presumes that his ancient “Elongated African” stock — a monolithic entity which, as we saw in part one, simply does not exist in the archaeological record — had kinky hair, he believes that the non-kinky hair texture that is common today among the Afro-Asiatic-speaking populations in the Horn is an indicator of exotic genetic influence. He hints that it may have been introduced in antiquity by Arabs, specifically. In reality, this non-spiralled hair form was the characteristic texture of the ancestral Cushitic speakers themselves. It was thus simply passed down from them to their modern descendants. We know this because, within the Horn region itself, every hunter-gatherer, Bantu and Nilotic population that has had extensive contact with early Cushitic groups has at least some members that possess soft-textured hair as a direct result of that interaction. Puccioni (1931) thus reports that, among the Ribi foragers of Somalia (who have significant Cushitic admixture), 60% of the individuals he examined had frizzy hair while only 40% possessed woolly hair. We also know this because the ancient peoples of the Kerma civilization in Sudan, barring a few assimilated individuals of Nilotic origin, uniformly had non-kinky hair. According to linguistic analysis by Marianne Bechaus-Gerst (1989, 2000), these Kerma folks spoke languages from the Cushitic branch of the Afro-Asiatic family. The pioneering Prussian Egyptologist, linguist and archaeologist Karl Richard Lepsius had the opportunity to examine the Kerma remains, and notes the following:
Speaking not as a trained anatomist but as one who has had the characteristics of different Nile Valley races pointed out to him by Prof. Elliot Smith and has handled the material for years, I may venture to add that the bones are not those of Negroes. A few cases of prognathous skulls occur and even, in the case of a woman in grave Kerma, tightly curled black negro wool on the head; but most of the men, especially the principal skeletons, had fine heads with straight black hair.
(For more details, see Did the ancient Cushites have soft-textured hair?.)
-

-
Somali politician Asad Osman Abdullahi (Majerteen Darod clan), representing a “pure” Cushitic physiognomy. The ancient Cushitic settlers of the Great Lakes region were described as “tall, bearded, long-haired, and ‘red’ in colour,” and thus would likely have had a similar phenotype. However, their great height surpassed that of most other Afro-Asiatic-speaking populations, except for the Tuareg Berbers.
-

-
Somali entrepreneur Hussein Adan Igeh (Isaac clan). Forensic analysis of ancient Cushitic specimens indicates that they had soft-textured wavy/curly/straight hair, which their modern descendants in Northeast Africa have inherited.
-

-
A soft-textured, wavy lock of hair belonging to a Kerma period individual excavated at Kadruka, Upper Nubia (northern Sudan). Ancient DNA analysis by Wang et al. (2022) has revealed that this specimen is genetically indistinguishable from the Cushites of the Early Pastoral Neolithic. (For more details, see Did the ancient Cushites have soft-textured hair?.)
-

-
An Iraqw boy (hunter-gatherer admixed Cushite). The Iraqw inhabiting Tanzania are among the last remaining descendants of the ancient South Cushitic-speaking settlers of the Great Lakes region. In the Iraqw community today, there are many individuals who are of assimilated Niger-Congo/Nilo-Saharan/hunter-gatherer origin. However, the Iraqw proper have managed to retain substantial Cushitic ancestry. This is particularly evident in terms of hair form since, although they mixed with their Tanzanian neighbors, certain Iraqw individuals still have the non-kinky hair texture of their Cushitic ancestors.
-

-
A Yaaku man (hunter-gatherer-admixed Cushite). The Yaaku, numbering just three individuals, are also among the last remaining descendants of the Great Lakes region’s early Cushitic settlers. Despite prolonged admixture with neighboring foragers, some Yaaku have likewise managed to retain the original soft-textured curly/wavy/straight hair of their Cushitic ancestors. This sets them apart from the Tutsi-Hima and Maasai, two Cushitic-admixed lacustrine groups whose exclusively kinky hair texture betrays their Bantu and Nilotic origins.
Additionally, Hiernaux indicates that the ancient C-Group pastoralists of Lower Nubia entered the Nile Valley from the Sahara and also settled in the Horn. He bases this on similarities in material culture and rock art. For example, John Desmond Clark asserts that “cattle and caprids (meaning here sheep/goat) were introduced to the lowlands of the Horn and the fringes of the Ethiopian plateau by C-group pastoralists from the Nile Valley and the surrounding arid regions” (cf. Barnett (1999)). Arkell (1961), moreover, identified pottery and stone work in Agordat, Eritrea, akin to that produced by the C-Group peoples (Fage and Oliver (1970)). Reisner (1912) likewise states that the “black- and brown-polished wares generally with incised white-filled patterns, peculiar to the Early Predynastic, Early Dynastic and the C-group periods[…] this pottery may be made in imitation of ebony or gourd vessels. The C-group patterns are suggestive of modern Somali, Sudanese and Nubian basketwork patterns, especially in the coloured examples.” Similarly, Stuart Munro-Hay writes that “pottery studies have suggested that the pre-Aksumite culture might owe something to Nile valley influences, specifically to the C-group/Kerma cultures (see O’Connor, Ch. 34, this volume), and in rather later times to Meroe/Alodia, as well as to contacts across the Red Sea on the Yemeni and Saudi coasts” (cf. Andah et al. (2014)). Furthermore, rock art at Dhawaale in northwestern Somalia, Harurona in the Wolayta region of Ethiopia, Abka in Sudan, and other archaeological sites in Northeast Africa include cross-in-circle motifs (also known as the encircled cross or cross-filled circle) (Bachechi (2005)). Istituto per l’Oriente (1979) notes that these are common symbols in C-Group art, which still feature prominently in Egyptian Coptic magic.
-

-
Cave painting at the Dhawaale site in northwestern Somalia featuring a cross-in-circle design. This artistic pattern has been found on pottery belonging to the C-Group culture, an ancient Afro-Asiatic-speaking population of Nubia.
-

-
Other examples of cross-filled circle symbols (#4-6), which have been observed on rock art at Harurona in Ethiopia and Abka in Sudan, as well as on material objects belonging to the C-Group culture. This symbol also still features prominently in Egyptian Coptic magic (Bachechi (2005)).
-

-
Rock art at the Dhawaale site in northwestern Somalia. The symbol in the center with looping edges resembles C-Group designs, further corroborating ties with this ancient culture.
-

-
An inverted pottery bowl belonging to the C-Group culture of Nubia. Symbols with looping edges are recurring motifs in C-Group art (Nubian Museum, Egypt).
Osteological affinities between the modern Afro-Asiatic-speaking groups of the Horn and the C-Group folks have also been established (see Bräuer (1980a) above; also Batrawi (1946)). According to Peter Behrens (1981) and Marianne Bechaus-Gerst (2000), the C-Group makers spoke languages from the Berber branch of the Afro-Asiatic family (this linguistic association is backed by the recent finding in Somalia of ancient Libyco-Berber/Tifinagh inscriptions, which were presumably left by early C-Group settlers). Like the coeval Kerma inhabitants, the C-Group folks were also found to have had non-kinky hair. In 1914, the Peabody Museum scholar Oric Bates led excavations in the Libyan/Western Desert, where he observed:
The so-called “C Group” cemeteries of Nubia, it was early recognized, were those of a distinctly non-Egyptian people. They extend in time from about the end of the Vlth Dynasty to the XVIIIth Dynasty, although the lower date is one to be stated with some reserve[…] Only by exception is the hair woolly or “peppercorn-like” ; as a rule it is straight or wavy.
With respect to lip form, Hiernaux asserts that the Tutsi Bantus actually tend to have thicker lips than the Hutu Bantus:
Lip thickness provides a similar case: on an average the lips of the Tutsi are thicker than those of the Hutu. In most cases, however, they are not everted as in many West Africans.
Finally, analysis of fingerprint distribution patterns around the world indicates that the Tutsi-Hima Bantu and Maasai Nilote inhabitants of the Great Lakes region share similar dermatoglyphic characteristics as other Niger-Congo, Nilo-Saharan and Khoisan-speaking groups. All of these populations have a low pattern index (PI) or pattern intensity index (PII) of under 12.00. Contrarily, the Afro-Asiatic-speaking populations of the Horn region have relatively high fingerprint pattern values of around 13.50 to 14.25, like other Afro-Asiatic speakers in North Africa and the Middle East (Rife (1953)). Of these groups, Yohannes and Bekele (2015) report that their Cushitic-speaking Oromo sample from Ethiopia has the highest pattern intensity index, valued at 15.01.

Global distribution of fingerprint (dermatoglyphic) pattern values. The Tutsi-Hima Bantus, Maasai Nilotes and other Niger-Congo, Nilo-Saharan and Khoisan-speaking populations are characterized by a low pattern index (PI) or pattern intensity index (PII) below 12.00. By contrast, the Afro-Asiatic-speaking populations in the Horn of Africa, North Africa and the Middle East have a relatively high pattern index of around 13.50 to 14.25 (Rife (1953)).
In summary, Hiernaux was gravely mistaken about the somatic traits of the ancient Caucasoids in East Africa. They were of a lighter complexion than he had envisioned, and they left behind a corresponding skin pigmentation allele to show for it. Since this mutation originated only a few thousand years ago in or near West Asia, these early Afro-Asiatic speakers could not have settled in East Africa earlier than the Neolithic. Likewise, genetic examination of associated human remains has established that these pastoralists did indeed ultimately arrive from North Africa and/or the Middle East. They also had non-kinky hair texture, quite unlike the spiraled hair that Hiernaux had envisaged for them. These vanished “Hamitic” peoples left a low-to-moderate imprint on various early Bantu and Nilotic groups that they encountered in the Great Lakes region. However, their actual Afro-Asiatic-speaking relatives can still be found to the north in the Horn.
Anthropometry
The core of Hiernaux’s theory is centered on a handful of external physical measurements on living subjects, which he argues establish an ancestral tie between his pure “Elongated Africans” (Tutsi-Hima, Maasai and Ful) and the Afro-Asiatic-speaking populations of the Horn (Beja, Galla/Oromo, Somali, etc.). Numbering 11 variables in total, these anthropometric means include stature, head length, head breadth, face height, face breadth, nose height, nose breadth, relative trunk length, cephalic index, facial index and nasal index.
Hiernaux has two Somali samples in his dataset, neither of which he measured himself. The first is a small sample of southern Somali individuals, “mostly of Sab descent”. These Sab Somali measurements were collected by the Italian anthropologist Nella Puccioni during an officially commissioned study published in 1917. The second, “larger sample of northern Somali belong[s] to various groups, the best represented being the Warsingali”. Hiernaux credits Lawrence Oschinsky (1954) with having gathered the Warsingali Somali measurements, although Oschinsky himself actually indicates that he obtained the data from “The Mediterranean Race in East Africa”, a chapter in Carleton Coon’s influential 1939 work The Races of Europe.
Hiernaux suggests that the two Somali samples manifest substantial anthropometric differences. Specifically, he contends that the southern Somali cohort bears a close similarity with his Tutsi sample from Rwanda (a sample drawn from an early 1954 study of his). Hiernaux also posits that the northern Somalis “are strongly Arabicized”, based mainly on their shorter stature and more narrow nose and face. In doing so, he makes several important errors.
First, Hiernaux somehow overlooks or ignores one major specification that Puccioni provides in his original description of the Sab collection. Puccioni explicitly states therein that the southern Sab cohort has greater Negroid influence than the other Somali groups.
As the historian I. M. Lewis notes:
Within the Ethiopic group, the Somali belong to the eastern division, and show very few Negroid characteristics as compared with the western division, which is quite notably Negroid. As a result of his very extensive examination of Somali physical types, Puccioni considers that the southern Sab confederacies show a higher degree of Negroid influence, corresponding to their part Negroid origin.
On the Sab, Lewis points out elsewhere that:
Linguistically the speech of the Sab differs from that of the northern pastoralists by about as much as French does from Italian. The gulf in language is thus much wider than that between any of the northern pastoral dialects. The distinction has also a strong historical component, for, as I shall presently show, the Sab are a conglomerate people, an amalgam of many different Somali groups with Galla and negroid elements. And where they differ from the northern pastoral clan-families their distinctive culture and social institutions reflect these mixed origins.
This would explain why the Sab are closer to the Tutsis than are the northern Somalis. That is, the higher Negroid element among the Sab is bringing them nearer to the Tutsis than otherwise would have been the case. The Tutsis are likewise drawn toward the Sab due to their greater Cushitic element than other Bantus.
Similarly, Hiernaux apparently ignores Coon’s detailed analysis of his northern Somali sample. This too is unfortunate because Coon stresses the group’s overall “Mediterranean Caucasoid” affinities:
There can be no doubt that the tall stature of the Gallas, Somalis, and Agaus is an old Hamitic trait, since both the negroid Sidamos and the Semites of Hadhramaut origin are much shorter. The tallness of this East African Mediterranean strain stands in contrast to the moderate stature of the Mediterranean Arabs across the Red Sea, and constitutes a characteristic difference between them. The bodily build of the East African Hamites is typically Mediterranean in the ratio of arms, legs, and trunk, but the special attenuation of the extremities among the Somalis is a strong local feature, which finds its closest parallels outside the white racial group, in southern India and in Australia.
Second, Hiernaux does not offer any comparative explanation to support his claims, much less multivariate analysis. He just takes it for granted that his 11 anthropometric measurements alone are similar enough to substantiate these assertions. But are they really?


Mean stature figures for the Sab (Dighil and Rahanuìn) and other Somali clans (Puccioni (1931)). *N.B. Puccioni’s stature average for the Dir includes Garre individuals. When measured alone, Boughey (1971) observed that the Dir proper are instead primarily tall, with a mean height of 172.9 cm/1729 mm.
A closer look at Hiernaux’s stature figures shows that the Sab, whom he indicates are the tallest Somalis, are three centimeters or a little over an inch shorter than the Tutsis (173 cm vs. 176 cm). The Warsingali Somalis and Galla are a full eight centimeters/three inches (168 cm vs. 176 cm) and five centimeters/two inches (171 cm vs. 176 cm), respectively, shorter than the Tutsi. Likewise, with respect to the facial index, the Sab Somalis are mesoproscopic or have a medium face type (88.5), whereas the Tutsi are leptoproscopic or have a long narrow face type (92.8). The Warsingali Somalis are, by contrast, almost hyper-leptoproscopic (94.1). As it is, these values don’t particularly lend strong support to Hiernaux’s contention that the Tutsi anthropometrically parallel the Sab. Worse, Hiernaux appears to outright misrepresent Puccioni’s original height data on the Sab (see Table 108 and chart above). Puccioni (1931) actually lists two mean stature figures for the Sab individuals that he examined: Digil (Dighil) males at 1677 mm/167.7 cm and Rahanweyn (Rahanuìn) males at 1698 mm/169.8 cm. These values make the Sab on average the shortest of the Somali clans, thereby directly undermining Hiernaux’s claims.
Franco et al. (2013) provide helpful visual guides on these anthropometric variables:

 Moreover, although Hiernaux emphasizes that the Tutsi’s and the Maasai’s relatively low nasal index is a key diagnostic feature that brings them closer to Afro-Asiatic-speaking groups than other Bantu and Nilotic populations, he completely ignores why those values are low to begin with.
Moreover, although Hiernaux emphasizes that the Tutsi’s and the Maasai’s relatively low nasal index is a key diagnostic feature that brings them closer to Afro-Asiatic-speaking groups than other Bantu and Nilotic populations, he completely ignores why those values are low to begin with.
 The nasal index is calculated by dividing the nasal width by the nasal height, and then multiplying that quotient by 100. From this equation, it is clear that a marked nasal height alone is sufficient to produce a low nasal index figure, even if the nasal width is not particularly low. This is, in fact, the situation with the Tutsi and Maasai. Unlike what Hiernaux insinuates, neither population has a nose breadth within the low range of the Afro-Asiatic-speaking populations (lowest for the Warsingali Somali at 34 mm). Both instead have the same moderate nasal width as the Ful sample from the northwestern region of the Central African Republic (39 mm). However, because of their unusually long noses, the Tutsi and Maasai still wind up with lower nasal indices than the Ful (69.5 for Tutsi, 72 for Maasai and 79.7 for Ful). Here too, though, their average values remain distant from that of the Warsingali Somali, who have a leptorrhine nasal index of 66 — squarely within the narrow Caucasoid range. Radlauer (1914) observed a similar leptorrhine nasal index of 65.7 in a general northern Somali (Hashiya) sample, so the Warsingali average is close to the modal range for northern Somalis as a whole.
The nasal index is calculated by dividing the nasal width by the nasal height, and then multiplying that quotient by 100. From this equation, it is clear that a marked nasal height alone is sufficient to produce a low nasal index figure, even if the nasal width is not particularly low. This is, in fact, the situation with the Tutsi and Maasai. Unlike what Hiernaux insinuates, neither population has a nose breadth within the low range of the Afro-Asiatic-speaking populations (lowest for the Warsingali Somali at 34 mm). Both instead have the same moderate nasal width as the Ful sample from the northwestern region of the Central African Republic (39 mm). However, because of their unusually long noses, the Tutsi and Maasai still wind up with lower nasal indices than the Ful (69.5 for Tutsi, 72 for Maasai and 79.7 for Ful). Here too, though, their average values remain distant from that of the Warsingali Somali, who have a leptorrhine nasal index of 66 — squarely within the narrow Caucasoid range. Radlauer (1914) observed a similar leptorrhine nasal index of 65.7 in a general northern Somali (Hashiya) sample, so the Warsingali average is close to the modal range for northern Somalis as a whole.
Oladipo et al. (2011) explain this standard pattern:
Nasal index measurement can be utilized in the analysis and classification of fossil remains as well as the study of living populations (Alex et al., 1996). Studies have shown that the Negroid race mainly of African descent have the platyrrhine nose type (Carleton, 1989).
[…]Oladipo et al. (2007) conducted a study on the morphometric analysis of the nasal parameters of Igbo, Ijaw and Yoruba ethnic groups of Southern Nigeria. The results obtained showed that an average Igbo had a mean nasal index of 94.1±0.37, Yoruba 89.2±0.30 and Ijaw 96.37±1.06. Thus the Ijaws had a significant higher nasal index (p<0.05). Fawehinmi et al. (2008) reported a mean nasal index of 98.5±0.93 and 94.1±1.18 for male and females of Kalabari ethnic group of Nigeria. The Somalia people in East Africa have a nasal index similar to that of European Caucasoid of 69.90 or less, which is of leptorrhine nose type (Porter et al., 2003; Carleton, 1989). The nasal index of African-American women is 79.70 Bantu speaking negreos and the bushmen of Africa as well as the Australoids of Australia are platyrrhine having a broad nose and a nasal index of 85.00 or more (Mulchand, 2004).

Nasal indices of Maasai individuals in Kenya. Most of the Maasai cohort have high nasal indices in the mesorrhine-platyrrhine (moderately broad) range, like other Great Lakes Nilotes. However, the presence in the sample set of leptorrhine (fine-featured) individuals of recent Cushitic origin — such as the “Mukogodo Maasai” i.e., assimilated Yaaku individuals from the Mukogodo forest — lowers the overall group average to a mesorrhine (moderate) nasal index (Leys and Joyce (1913)).
David Blackwell (1984) went further than Hiernaux’s basic nasal index calculations and photogrammetrically measured all aspects of the Tutsi Bantu’s and the Hehe Bantus’ nasal morphology. He found that there were no significant differences with respect to these Niger-Congo-speaking populations’ other nose-related indices, nor any marked disparities between them in terms of their nasal prominence, nasal bridge distance, columella length, nasal height, nasal angle, nasal wing/septum relationship, or nasal region vertical relationship. By measuring the nasal breadth index and inter-occular nasal width index, Blackwell was also able to gauge just how wide the average Tutsi’s nose is relative to his/her facial breadth. He found that while the Tutsi individuals he examined had a lower nasal index than his Hehe cohort, the sampled Tutsis in fact had a considerably larger nasal breadth index and inter-occular nasal width index than their Hehe counterparts. According to the scientist:
These results suggest that although the Hehe’s noses are wider in absolute terms the Tutsi’s are wider relative to their facial widths. The nasal index, which is nasal height divided by nasal breadth, was larger in the Hehe and since there was no significant difference in nasal heights this again suggests the Tutsi’s noses are wider relative to face width than the Hehe’s.
Note that although the Hehe are Bantu speakers, these anthropometric variables are also compatible with the idea that the Tutsi are of Nilotic origin since the coupling of a narrow head/face and a proportionately broader nose is a distinguishing characteristic of Nilotes (see discussion and anthropometric table below under Ecology, rock art and genetics).
-

-
A Tutsi man and woman. Because the Tutsis’ Bantu ancestors in the past assimilated some Southern Cushites, the average Tutsi individual today tends to have narrower facial features than other Niger-Congo/Nilo-Saharan speakers. However, the nasal width of the Tutsi (estimated at 39 mm, similar to the Maasai Nilotes) remains significantly broader than that of Afro-Asiatic speakers. Likewise, photogrammatic analysis of the Tutsi Bantus’ nasal morphology as compared to that of the Hehe Bantus indicates that these Niger-Congo-speaking populations are similar to each other on most indices, including nasal prominence, nasal bridge distance, columella length, nasal height, nasal angle, nasal wing/septum relationship, and nasal region vertical relationship. Moreover, although the Tutsis generally have a lower nasal index than their Hehe brethren, the Tutsi also have a considerably larger nasal breadth index and inter-occular nasal width index. This “again suggests the Tutsi’s noses are wider relative to face width than the Hehe’s” (Blackwell (1984)).
As we saw in part one, the tall height of the Tutsis, as well as their long heads and less broad facial features than other Bantus, is due to contact with the Southern Cushitic “giants” of yore, whom the Tutsis’ ancestors assimilated. These physical attributes were not obtained from a mythical “Elongated African” entity, as Hiernaux muses. After having been introduced, these traits were, per the French social geographer Dominique Franche, maintained via selective breeding (though diet may have also played a part). Through this process, the Tutsis’ Bantu forebears would choose mates according to how closely those individuals conformed to the new Cushitic-influenced beauty ideals. This is evident given the antiquity and low-to-moderate level of Cushitic gene flow into the Tutsi population, which on its own would not have been enough to substantially modify that community’s ancestral Bantu morphology. Prior to said Cushitic admixture, the Iron Age inhabitants of Rwanda all had a physiognomy like their Niger-Congo-speaking relatives in West Africa, and most still do (cf. Rightmire (1975); Hiernaux (1960)). With regard to the Maasai Nilotes, who it has been established also have some Southern Cushitic influence, their shorter height relative to the Tutsi is largely due to the fact that they more intensively interbred with diminutive hunter-gatherer groups (cf. Dobon et al. (2015)).
(For more details, see Were the ancient Cushites slender?.)
Later anthropometric and craniometric studies
-

-
A Sahrawi Berber (Moor) man with the beidan phenotype. This “Mediterranean Caucasoid” physiognomy is characterized by a light brown to olive complexion. It is most common today among Berbers and Copts, who are the groups in North Africa that were least impacted by foreign intermixture. Closely corresponds with the original “brun-type” of the ancient Libyans, the predynastic Egyptians and early Cushites, as depicted on ancient murals and engravings in the Nile Valley. Analogous with the cad phenotype found amongst the Cushitic-speaking populations in the Horn of Africa.
-

-
A Sahrawi Berber/Moor man with the amrani phenotype. Essentially of the same ancestral stock as the beidan North Africans, albeit with minor Sub-Saharan admixture. The amrani physiognomy can be found throughout North Africa but is most common in the southern areas, particularly among Saharan Berbers (Sahrawi Moors, Tuareg & Siwa Berbers), southern Moroccans, northern Sudanese and southern Egyptians. It is roughly equivalent with the mariin phenotype that is widespread among the Afro-Asiatic-speaking populations in the Horn region.
-

-
A Sahrawi Berber Moor man with greater Sub-Saharan African admixture. All contemporary Maghrebis carry some degree of Sub-Saharan African admixture, which primarily consists of ancient SSA admixture embedded within their Iberomaurusian component and recent SSA admixture derived from interaction with Niger-Congo-speaking groups of West Africa. In his phenotype, this Sahrawi individual shows marked traces of these Sub-Saharan African elements, mainly in terms of hair form. As such, he represents the Maghrebi equivalent of the yusuur physiognomy of the southern part of the Horn and the northern half of the Great Lakes region, a broad area where Afro-Asiatic speakers come into contact with various Sub-Saharan African populations.
After the publication of Hiernaux’s The People of Africa, a number of studies were independently released, which contradicted, invalidated or otherwise corrected his “Elongated African” theory. The earliest of these works was Hiernaux’s own “Afrique moyenne” published the following year, in 1975. It was part of the Afrika ein volume in Rassengeschichte der Menschheit, a series on human diversity overseen by the anthropologist Eugene Strouhal. Based on anthropometric means, Hiernaux calculated the biological distance between hundreds of populations from all major ethnolinguistic groups below the Sahara. He included the Moors and Beja in his multivariate analysis, two populations that he had mentioned in passing in his book but did not actually examine. Hiernaux found that, after the Mbuti Pygmies (who, on account of their diminutive height, were obviously very divergent), the Afro-Asiatic-speaking groups on the northern fringes were the most distant. Of these latter populations, the Warsingali Somali were the furthest at a distance of 981, followed closely by the Bisharin subgroup of the Beja at 977 and the Moors at an average of 931. The Warsingali Somalis and Moors also formed a “constellation” unto themselves. Of the Bantu and Nilotic populations, the Tutsis of Burundi and Rwanda at averages of 859 and 829, respectively, were the nearest to the Afro-Asiatic-speaking groups. This is to be expected given their Cushitic admixture. Hiernaux, however, went further and suggested that this demonstrated that the Tutsi were not of Bantu origin but were instead settlers from the Galla area in southern Ethiopia, who later mixed with Bantu agriculturalists in the Great Lakes region and adopted their language — basically, a return to his initial Hamitic origin theory.
In 1976, G. P. Rightmire set out to evaluate the strength of Hiernaux’s biodistance analysis. Using both cranial metric and non-metric multidimensional scaling on 61 populations in Sub-Saharan Africa, including the Maasai Nilotes, he found no evidence for an “Elongated African” ancestral stock. Rightmire instead confirmed that the Afro-Asiatic-speaking populations of Northeast Africa were biologically distinct from the Nilo-Saharan, Niger-Congo and Khoisan groups. Of all the samples examined, the scientist observed that the Warsingali Somali of northern Somalia were overall the most distant, with the Oromo/Galla of Ethiopia and the Sab Somali of southern Somalia following suit:
This departure from “typical” African frequencies has been confirmed for various marker genes among Ethiopians (Ikin and Mourant, ’62; Harrison et al. ’69), so that the present peripheral position of the Galla and related Cushitic speaking groups is not surprising.
Additionally, Rightmire noted that the Nilo-Saharan-speaking Kunama of Eritrea also fell into the Afro-Asiatic cluster, a situation ascribable to substantial gene flow from the neighboring Afro-Asiatic-speaking communities (for Y-DNA evidence of such genetic introgression, see Trombetta et al. (2015), Supplementary Table 7; 65% of Kunama individuals have been found to bear the Afro-Asiatic-affiliated E1b1b haplogroup). This affiliation with Afro-Asiatic speakers can, alternatively, be interpreted as supporting the Kunama’s claim that they are not of Nilotic origin, but instead Abyssinian descendants who adopted a Nilo-Saharan language (Elisée Reclus (1899) notes: “The Kunama, established in the country since time immemorial, claim to be immigrants of Abyssinian origin, and the Abyssinians themselves look upon them as descendants of the ancient Aksumites”). For their part, the Maasai Nilotes of the Great Lakes craniometrically grouped with the other Nilo-Saharan and Niger-Congo samples, consistent with their comparatively lower Cushitic admixture. However, the Maasai did evince some non-metric affinities with the Afro-Asiatic-speaking populations. This in itself does not mean much, though, since research has demonstrated that non-metric/discrete/epigenetic trait analysis, whether cranial or dental, is really only useful for intra-population or within-population studies. Wilson (2010) explicates:
The suite of nonmetric traits expressed on an individual’s skeleton and dentition has been assumed to be genetically inherited. In other words, it is assumed that the phenotype (observable characteristics) of an individual will provide direct information about his or her genotype (genetic constitution). This assumption has allowed many researchers to use nonmetric traits to assess genetic relatedness within and between populations in the archaeological record (Matsumura, 2007). Understanding these relationships in past populations (especially those without written histories) can provide information about migration patterns, residence patterns, population structures, and human origins and evolution (Hanihara, Ishida & Dodo, 2003; Hlusko, 2004; McLellan & Finnegan, 1990; Lane & Sublett, 1972; Turan-Ozdemir & Sendemir, 2006). The term “biodistance” is commonly used to describe genetic relatedness. Saunders and Rainey (2008) describe biodistance as a measure of the amount of divergence; less divergence is equal to a closer genetic relationship (Saunders & Rainey, 2008; Sherwood, Duren, Demerath, Czerwinski, Siervogel, & Towne 2008). It should be noted that research has shown nonmetric traits to be population specific and therefore only really useful for intrapopulation analyses (Cheverud & Buikstra, 1981; Kohn, 1991).
When utilized in an inter-population or between-population context, non-metric trait analysis instead often results in spurious biological groupings, which contradict data collected through other, more scientifically robust methods. Sarfo (2014) thus remarks:
Epigenetic traits of the human skeleton, particularly of the skull, have had a long history in the study of paleogenetics. The last century, in particular, witnessed both the rise and fall of the use of epigenetic nonmetric traits in paleopopulation research. Part of the fall was attributed to rise of molecular anthropology in the 1980s and 1990s, whereby actual DNA could be analyzed to determine genetic relationships. This was coupled with problems in the research designs of the traditional morphologically-based paleopopulation genetics and the misunderstanding of the expression of epigenetic traits in the development of morphological variation. The pioneering article by Berry and Berry (1967) posited that nonmetric epigenetic cranial traits were highly genetic, were independent of each other, were independent of age and sex, and could be easily scored and standardized. Critical evaluation of these assumptions in the latter decades of the 20th century resulted in many challenges, particularly in the development of proper trait lists (Ossenberg 1976). For example, of the original 30 traits overviewed by Berry and Berry, only 7 can be confidently used to study paleopopulation genetics (Molto personal communication 2014). Yet, the vast majority of researchers in the period between the 1970s and 1990s utilized the Berry and Berry trait list with conflicting results arising as a consequence. Researchers who examined the assumptions generally found that many nonmetric traits were not independent of age, sex and symmetry (Ossenberg 1969, Suchey 1975, Molto 1985), and above all were not easily scored and standardized (Molto 1983). The latter is a fundamental requirement of the scientific method.
Eckhardt (1989) summarizes the problem with non-metric analysis in inter-population/between population studies as follows:
As noted in preceding sections of this review, there are reasonable prospects that better data on specific modes of inheritance of various skeletal traits, or at least estimates of heritabilities and the numbers of genes contributing to multifactorial characteristics, can help us to resolve one current paradox in studies of population affinity: that sometimes nonmetric traits suggest different patterns of relationship from those based on other data, including metric skeletal traits (Corrucini 1974). According to Brothwell (1959) nonmetric traits showed Chinese and Peruvian crania to be more closely related than Anglo-Saxon and London crania; in the same study Chinese were grouped more closely to North American Indians than Anglo-Saxons were to recent Germans or Melanesians to Polynesians. In another investigation Berry & Berry (1967) got results that placed South American Indians closer to Burmese and even to Africans than to North American Indians; also, Egyptian crania were placed nearer to those from South America than to a population from Palestine. In the other direction, Finnegan (1972) could not differentiate Florida Indians from several groups of Northwest Coast Indians.
Given the caveats above, it is hardly surprising that non-metric cranial and dental analyses of the Horn region’s Afro-Asiatic-speaking populations have shown little consistency. The conclusions of these studies have instead proven dubious, being at odds with cranial/dental metric and genetic analyses conducted by other researchers. For instance, Rightmire (1976) noted “Mediterranean” cranial non-metric affinities for his Cushitic samples (Warsangali Somali, Saba Somali, Oromo/Galla), whereas Hanihara (2003) reported that his sample of Ogaden Somali individuals (i.e. Ethiopian Somali) showed Sub-Saharan cranial non-metric ties. We can be confident that this is just another example of non-metric analysis going awry, as Vicente et al. (2019) genomically observed that their Somali North sample (Ethiopian Somali) actually has less Sub-Saharan admixture than their Somali South sample (Sab Somali). Likewise, Haddow (2012) analysed dental non-metric traits and found that their contemporary Ethiopia sample had closest biodistances to their ancient and modern North Africa samples, while Irish et al. (2007)‘s Tigrayan sample from Ethiopia/Eritrea instead shared nearest dental non-metric similitude with the ancient Jebel Moya sample from Sudan. We can be reasonably certain that this, too, is incorrect since Mukherjee (1955) indicates that the Tigrayan individuals he examined craniometrically cluster with the ancient Afro-Asiatic-speaking samples from North Africa; his Jebel Moya cohort, in contrast, craniometrically groups with the Sub-Saharan samples.
Moreover, recent genome research has identified specific alleles that are associated with craniofacial development (e.g. the EDAR, DCHS2, RUNX2 and GLI3 genes). These genetic findings have substantiated the validity of cranial/dental metric analysis for both intra-population and inter-population studies (Roosenboom et al. (2016); Adel et al. (2021)).
-
![Cranial metric analysis of Afro-Asiatic, Nilo-Saharan, Niger-Congo and Khoisan-speaking populations in Sub-Saharan Africa. The Afro-Asiatic-speaking Warsingali Somali, Oromo and Sab Somali samples cluster separately from the Sub-Saharan samples (including the Cushitic-admixed Maasai Nilotes), owing to ancestral ties with "Mediterranean Caucasoids." As Rightmire notes, "the departure from typical "African" frequencies has been confirmed for various marker genes among Ethiopians[...] so that the present peripheral position of the Galla and related Cushitic speaking groups is not surprising." The Kunama of Eritrea also fall in the Afro-Asiatic cluster due to significant gene flow from neighboring Cushitic and Ethiosemitic-speaking populations. This is the standard clustering pattern for these samples in metric analysis (both cranial and dental), which research has established is genetically inherited (Rightmire (1976)).](https://landofpunt.files.wordpress.com/2015/09/rightmire1976-fig2.png?w=264)
-
Cranial metric analysis of Afro-Asiatic, Nilo-Saharan, Niger-Congo and Khoisan-speaking populations in Sub-Saharan Africa. The Afro-Asiatic-speaking Warsingali Somali, Oromo and Sab Somali samples cluster separately from the Sub-Saharan samples (including the Cushitic-admixed Maasai Nilotes), owing to ancestral ties with “Mediterranean Caucasoids.” As Rightmire notes, “the departure from typical “African” frequencies has been confirmed for various marker genes among Ethiopians[…] so that the present peripheral position of the Galla and related Cushitic speaking groups is not surprising.” The Kunama of Eritrea also fall in the Afro-Asiatic cluster due to significant gene flow from neighboring Cushitic and Ethiosemitic-speaking populations. This is the standard clustering pattern for these samples in metric analysis (both cranial and dental), which research has established is genetically inherited (Rightmire (1976)).
-

-
Cranial non-metric analysis of Afro-Asiatic, Nilo-Saharan, Niger-Congo and Khoisan-speaking populations in Sub-Saharan Africa. The Afro-Asiatic-speaking Warsingali Somali, Oromo and Sab Somali samples as well as the Kunama of Eritrea again cluster separately from the Sub-Saharan samples. However, the Maasai Nilotes of the Great Lakes region this time show affinities with the Afro-Asiatic cluster. This is a reflection of the inconsistent nature of non-metric analysis (both cranial and dental), which research has shown is not genetically stable but rather varies based on symmetry, sex and age (Rightmire (1976)).
-

-
Dental metric analysis of ancient and modern global populations. The Somalia sample (Ogaden Somali) has a similar overall tooth size (microdont/mesodont) as the ancient Egyptian (Predynastic & Dynastic) and Middle Eastern samples. By contrast, the modern Kenya (Kikuyu Bantus & Turkana Nilotes) and Tanzania (Haya) samples have a large tooth size (megadont) like the Southern African and West African samples (Hanihara and Ishida (2005)). This concurs with Brace (1993), who notes that “most of our sub-Saharan samples fall into the “megadont” category used by Flower to indicate relative tooth size[…] but the Somalis from the Horn of East Africa sit right on the dividing line between “mesodont” and “microdont.”
-
-
Dental non-metric analysis of ancient and modern African populations. The Ethiopia sample has closest biodistance values with the North African samples, particularly the Carthaginian (0.000), Badarian Egyptian (0.001), Pharaonic Nubian (0.002), Kabyle Berber (0.003), and A-Group Nubian (0.005) samples. Other Horn of Africa and North Africa samples in non-metric analyses have instead shown Sub-Saharan ties, which reflects the spurious nature of non-metric analysis as compared to the consistent and genetically-controlled metric analysis (Haddow (2012)).
-

-
Dental metric analysis of the dentition of South Cushitic-speaking Iraqw, Khoisan-speaking Hadza, and Nilo-Saharan-speaking Datoga individuals of Tanzania as compared to that of Caucasians and American Negroes. Although they have interbred with neighboring Bantu/Nilotic/hunter-gatherer populations, the Iraqw still group with the Caucasian sample (Ikeda and Hayama (1982)). This clustering pattern concurs with Gerhard Heberer (1963)’s observation that the early Cushitic settlers of the Great Lakes region were “Mediterranean Caucasoids with Caucasoid teeth.”
-
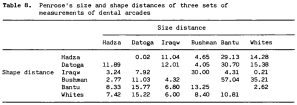
-
Dental metric analysis of the arcades of Iraqw, Hadza, Bushman, Datoga, Bantu and White individuals. The Iraqw sample has nearest biodistance to the Whites sample (0.21), for “the findings in measurements of dentition and dental arch suggest only close affinities of the Iraqw to the Whites” (Ikeda and Hayama (1982)). This again highlights the fact that the Iraqw have managed to retain significant Cushitic ancestry, as also indicated by genome (Scheinfeldt et al. (2019)) and haplogroup (Wood et al. (2005)) analyses.
-

-
Cranial non-metric analysis of ancient and modern global populations. Some “rather odd values in Table 4 are the greater likeness of the Egyptians to the Ashanti of Nigeria than to the inhabitants of Lachish in Palestine, and the lack of distinctiveness between West Africans and North Indians.” Such biologically incongruent groupings are typical of cranial and dental non-metric (discrete/epigenetic) analysis. Cranial and dental metric analysis, on the other hand, serves as a reliable measure of biological inheritance (Berry and Berry (1967)).
-
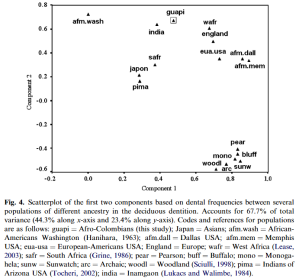
-
Dental non-metric analysis of global populations. The South Africa sample clusters with the Japan sample, the Afro-Columbian sample clusters with the India sample, the African-American samples from Dallas and Memphis cluster with the European American sample, the African-American sample from Washington clusters instead with no other population, and the West Africa sample clusters with the England sample. Such nonsensical results are commonplace in non-metric analyses, whether dental or cranial (Delgado-Burbano (2007)).
All in all, Rightmire attributes the morphological distinctiveness of the Afro-Asiatic speakers of the Horn of Africa to shared ancestral ties with “Mediterranean Caucasoids.” On this point, the linguist and anthropologist Harold C. Fleming remarks that Hiernaux’s peculiar perspective was likely shaped by his specialization in Central Africa, where the presence of any exotic influences among the local Niger-Congo-speaking populations is uncommon. Among the Afro-Asiatic-speaking groups in the Horn region, on the other hand (Fleming’s own area of focus), such “Mediterranean” traits are the norm. Fleming writes:
The most recent study of physical classification in Africa, GP Rightmire (1976), corrected and expanded Jean Hiernaux’s massive attempt to classify sub-Saharans. Rightmire used sophisticated measures of genetic distance and multidimensional scaling on the problem, but still kept his focus on black Africa. He however found that his Nilotes (Nuer, Alur, Kakwa, Luo, Masai) were distant from his northeast Africans (Sab, Somali and Galla) and gently suggested that Garn’s East Africans really meant northeast Africans rather than Nilotes. Hiernaux had also lumped Nilotes with Ethiopians apparently because of the linearity common to both — sometimes. I will take it as established then that Rightmire’s work has corrected Garn and Hiernaux with respect to southern Sudanese differences from Ethiopians. It is also clear, as a metaphysical matter, that Africanists who do physical classifications look at Ethiopia from the vantage point of the Congo and rarely consider “Mediterranean admixture” in terms other than the dilution of a basically African population. This viewpoint can come as a great shock to an Ethiopianist, accustomed to the vista from Bablyon.

An Ababda Beja girl, retaining the fine-features, facial orthognathism and soft-textured hair of her Cushitic Blemmyes (Bougaioi) ancestors.
In 1979, the anthropologists Strouhal and Jungwirth conducted a study, which compared the cranial affinities of modern East African “Negroes” from Rwanda and Burundi against those of late Roman-early Byzantine period skeletons excavated at Sayala, Egyptian Nubia. The Sayala specimens all belonged to the Pan-Grave culture and were exhumed from five cemeteries at the site (three cemeteries labeled CI-III, the N cemetery, and the A cemetery). Strouhal and Jungwirth report that most of the individuals from the CI-III and N burial complexes were of “Europoid” physical type (84.5%), whereas the individuals interred at the A cemetery were primarily of mixed “Europoid-Negroid” physiognomy (57.1%). The scientists also cross-analysed the discrete (non-metric) cranial traits of the CI-III cemetery Pan-Grave specimens with those of their Rwanda and Burundi “Negro” samples, and concluded that there was no affinity. This finding once more underlines the non-Cushitic origins of the Tutsi-Hima Bantus since the “Europoid” Pan-Grave culture bearers of the CI-III cemetery — with whom the East African “Negroes” from Rwanda and Burundi share no morphological ties — are believed to have originally been of Cushitic ancestral stock. Specifically, Strouhal and Jungwirth assert that “the Sayala CI-III series would represent the Blemmyes, a population originating from the Eastern Desert.” An early Coptic inscription refers to the Blemmyes as the Bougaioi, an appellative that is rendered as Beya in a later text retrieved from Adulis. This is an obvious allusion to the Cushitic-speaking Beja people, who still inhabit the Eastern Desert like their Blemmyes ancestors at the CI-III cemetery in Sayala, Egyptian Nubia (cf. Winstedt (1909)). (*N.B. In due course, some Nilotic peoples would gradually have been assimilated into the Pan-Grave community. These outsiders appear to have contributed the alien “Negroid” and mixed “Europoid-Negroid” physical types that have also been observed at various Pan-Grave sites, opposite the original “Europoid” physical type of the Cushitic Blemmyes.)

Anthropometric analysis of various Afro-Asiatic, Niger-Congo, Nilo-Saharan and Khoisan-speaking samples. The unadmixed Cushitic speakers from Northeast Africa (Beni Amer Beja and Hadendoa/Amara Beja) are distinct from all of the examined Sub-Saharan African populations, sharing close affinities only with each other (1.09 biodistance). The mixed Cushitic sample from Southeast Africa (Iraqw) is nearest to the Datog Nilotes, due to documented intermarriages. The Tutsi Bantu individuals are not especially close to any population, though they do bear somewhat remote ties with the Datog Nilotes (3.06 biodistance). The Sandawe foragers of Tanzania are morphologically very similar to the adjacent Nyaturu Bantus. Of the hunter-gatherer/Nilo-Saharan/Niger-Congo populations, the Sandawe are also the least isolated from the unadmixed Cushitic samples (3.94 biodistance from the Beni Amer Beja). This is attributable to absorption of Cushitic pastoralists by the Tehla or herder division of the Sandawe (Ikeda and Hayama (1982)).
Ikeda and Hayama (1982) published an anthropometric analysis comparing the Tutsi Bantus, Hutu Bantus and other Niger-Congo-speaking populations with various Nilo-Saharan, Khoisan and Afro-Asiatic-speaking groups. The study is unique in that it included among its samples unadmixed Cushitic individuals from Northeast Africa (viz. Beni Amer Beja, Hadendoa Beja and Amara Beja), Cushitic individuals from Southeast Africa with significant Niger-Congo, Nilo-Saharan and hunter-gatherer admixtures (Iraqw), and Niger-Congo, Nilo-Saharan and hunter-gatherer individuals with significant Cushitic admixture (Tutsi, Datog and Sandawe, respectively). The unadmixed Cushitic groups were found to be distinct from all of the examined Sub-Saharan African populations, sharing a close relationship only with each other (1.09 biodistance between the Beni Amer and Hadendoa/Amara Beja samples). The mixed Iraqw from Tanzania were nearest to the Datog Nilotes, consistent with their documented intermarriages (0.77 biodistance). They also showed more remote ties with most of the other samples, which are in line with known population contacts. Interestingly, the Tutsi individuals were not particularly closely affiliated with any sample; their nearest association was with the Datog cohort, at a notable biodistance of 3.06. This was not the case for the Sandawe, who shared very close morphological affinities with the neighboring Nyaturu Bantus of northern Tanzania (0.41 biodistance). Of all the studied groups, the Sandawe were also the least isolated from the unadmixed Cushitic samples, at a biodistance of 3.94 from the Beni Amer Beja. This suggests that most of the Sandawe individuals in the dataset belonged to the Tehla, the herder division of the Sandawe forager community, which is known to have assimilated many Cushitic pastoralists (due to these past interactions, the Sandawe today have an average of ~30% Eurasian admixture; see Scheinfeldt et al. (2019)).

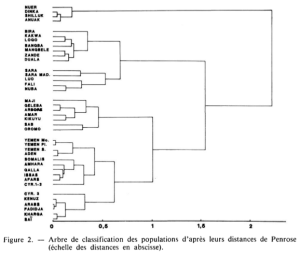
Anthropometric relations between ethnolinguistic groups in Africa and the Arabian peninsula. The Cushitic and Ethiosemitic-speaking populations in the Horn of Africa cluster with other Afro-Asiatic speakers in North Africa (northern Libyans and Egyptians) and the Arabian peninsula (Yemenis). Nubians also group with the Afro-Asiatic-speaking samples rather than with the Nilo-Saharan and Niger-Congo-speaking populations (Billy (1988)).
In 1988, G. Billy compared the morphological relationships of the Afro-Asiatic-speaking populations in the Horn with each other and with neighboring groups. He used the same northern Somali and southern Sab samples as Hiernaux did (taken from Coon and Puccioni, respectively). However, unlike Hiernaux, Billy did not restrict his samples to populations below the Sahara since he also examined other Afro-Asiatic speakers from North Africa and Southern Arabia. Running various multivariate analyses, which Hiernaux had likewise neglected to do in his book, Billy found that the Afro-Asiatic-speaking groups were all morphologically closest to each other. Furthermore, contrary to Hiernaux’s suggestion therein that the Somalis were anthropometrically intermediate between the Yemeni Arabs and the Tutsis and that the Galla were intermediate between the Somalis and the Tutsis, the Somali and Galla as well as the other northern Horn samples clustered nearest to all four Yemeni samples in the dataset in addition to the littoral Cyrenaican/northern Libyan and Egyptian samples, but not with any Bantu or Nilotic population. Unsurprisingly, given Puccioni’s caveat, the Sab sample grouped closest to populations inhabiting southern Ethiopia (mainly Omotic groups). This was due to what Billy described as a greater morphological tendency on their part toward broader noses and faces. However, the Sab were still closer overall to the other Afro-Asiatic-speaking populations.
Billy thus concluded that:
It appears that populations living at present in Egypt and Sudan are weakly influenced by the populations of Equatorial Africa but strongly related to coastal populations of Ethiopia and the arabian peninsula to a lesser extent.
The physician and anthropologist Alain Froment, the current Scientific Director of the anthropological collections at the Musée de l’Homme in Paris, would similarly highlight the morphological proximity between the Afro-Asiatic-speaking populations on either side of the Red Sea. Froment had worked closely with Hiernaux earlier in his career (they co-authored at least one publication in the 1970s), so he was quite familiar with his colleague’s theories. In a 1992 study, Froment, apparently unaware that Somalis and Gallas are Afro-Asiatic speakers with a different biohistory from Nilo-Saharan groups, lumped a Somali and Galla joint sample with a Nilotic Maasai one. Predictably, he found that his combined sample pulled towards Europe due to the Somali and Galla’s “Caucasoid” craniometric affinities, but also towards the Sub-Saharan African centroid on account of the presence of the Maasai Nilotes in the sample. By 1994, Froment had grown more familiar with the marked biological differences between the Afro-Asiatic-speaking populations in the Horn and Sahara versus their Bantu and Nilotic neighbors. In a retrospective paper on the ancient Egyptians published that year, he noted his joint Somali-Galla sample’s close craniometric ties with the Proto-Mediterraneans as well as with other Afro-Asiatic-speaking populations in North Africa and the Middle East. He also criticized Hiernaux’s consistent omission of North African samples from the latter’s various analyses. Froment quite rightly remarked that this oversight on the part of Hiernaux made it impossible to ascertain the actual morphological transitions between the Horn region and the Mediterranean Basin.
Additionally, during his The People of Africa phase, Hiernaux had argued that changes in morphology between populations inhabiting different regions below the Sahara were gradual and clinal. However, both Froment and W. W. Howells found evidence to the contrary in their multivariate analyses. They instead noted a sharp morphological discontinuity between the Afro-Asiatic-speaking populations in the Horn and the adjacent Bantu and Nilotic groups. I. M. Ribot writes:
Multivariate analyses of both Howells (1989) and Froment (1992a, b) agreed on the following points: in contrast to the observations of Hiernaux (1976), the variation within sub-Saharan Africa was not entirely continuous, as significant differences were observed between regions; and these marked geographical differences were well shown especially when using two cranial variables related to both bizygomatic breadth of face and nasal breadth.
[…]Furthermore, in agreement with Howells (1989) and Froment (1998) again, but in contrast to the observations of Hiernaux (1974), the range of variation in sub-Saharan Africa was not continuous or clinal, as significant differences were still observed between the different regions.

Craniometric relations between populations in Africa, the Middle East and Europe (Froment (1999)).
In 1999, Froment conducted another large-scale craniometric analysis for a paper on Sahelian populations. This time he included separate Somali-Galla and Sab samples, as well as Maasai, Tutsi, Ful (Peul), Berber, Nilotic and West African samples and Egyptian, Maghreban, Tuareg, Sub-Saharan, Middle Eastern and European centroids — 636 male and female global populations in total. Froment used six cranial measurements, including head length and head breadth, face length and face breadth, and nose length and nose breadth. He found that the Galla-Somali lumped sample was distant from the Maasai, Tutsi and Ful samples, and was instead nearest to the Middle Eastern and Bedouin/Lower Egyptian centroids. On the horizontal axis, where most of the variation between the samples was contained (77%), the Galla-Somali sample was again closest to the Bedouin/Lower Egyptian centroid in addition to the Sab Somalis, the Tuareg centroid, the Maghreban centroid, the Moors of the Western Sahara (the Sahrawi), and the Kel Kummer male Tuareg sample. The Sab Somali sample was, in turn, nearest to the Tuareg and Bedouin/Lower Egyptian centroids, while being closest to those samples on the key horizontal axis as well as to the Galla-Somali lumped sample, the Kel Kummer males and the Maghreb centroid.

Craniometric analysis of Afro-Asiatic, Niger-Congo, Nilo-Saharan and Khoisan populations. The Fulbe (Peul) and Tutsi samples are positioned on the margins of the Niger-Congo/Nilo-Saharan cluster, with a pull toward the separate Afro-Asiatic cluster. In the case of the Fulbe, this placement is primarily due to gene flow from ancient Berber and European populations. In the Tutsis’ case, this clustering pattern is attributable to both absorption of South Cushitic groups and sexual selection. Since the Tutsis experienced less overall gene flow from Afro-Asiatic-speaking populations than did the Fulbe, they would have had to have turned to careful mate selection in order to further alter their ancestral Bantu morphology (Froment (1998)).
In 2013, Terrazas Mata and Benavente craniometrically compared Tutsi, Hutu and other Bantu individuals from eastern Africa with various Niger-Congo, Nilo-Saharan, Khoisan and Afro-Asiatic-speaking populations. Their study is unique in that it includes a sample of an early Tutsi king, one Cyirima Rujugira (r. 1675-1708), borrowed from Ribot (2003). Hiernaux had previously analysed this ruler’s skeletal remains and reported that Cyirima shared affinities with the modern Tutsi (e.g. same average height of 176.539 cm) and to a lesser extent with the Hutu (cf. van Noten (1972)). Terrazas Mata and Benavente subsequently confirmed the close ties between the Tutsi king and Ribot’s other Bantu samples, lumping these related Niger-Congo-speaking individuals into a cohort labeled “Eastern Africa” (comprising King Cyirima and Rwandan Tutsi, general Burundi, Pare, Shambaa, Dschagga, Teita and Haya persons). The scientists found that this Eastern Africa Bantu sample clustered with two other Bantu-speaking groups from the Great Lakes region, the Hutu and Teita, “all known to be archaeologically and ethnographically similar and probably recently derived from a common ancestral population.” By contrast, their Horn of Africa lumped sample, consisting of Cushitic-speaking Somali, Beja and Oromo/Galla individuals, clustered instead with the Dynastic Egyptian sample. Terrazas Mata and Benevente indicate that this is because “these groups seem to represent migrations of groups from the Middle East, who spoke languages from the Afro-Asiatic family.”
Thus, contrary to Hiernaux’s suppositions, Terrazas Mata and Benavente, Froment, Billy and Rightmire had all empirically established that the Somali, Oromo/Galla and Beja are morphologically much closer to other Afro-Asiatic-speaking populations in North Africa and the Arabian Peninsula than to Nilo-Saharan and Niger-Congo-speaking populations, including groups with some Cushitic or Berber admixture like the Maasai, Tutsi and Ful.
Serology
Besides anthropometry, Hiernaux in his book The People of Africa relies on serology, or blood work, to support his “Elongated African” hypothesis. He focuses on relative differences in frequencies in two serological systems: the M allele in the MN system, and the R0 (cDe) allele in the Rh system. With regard to M, Hiernaux indicates that the average frequency for the allele in Sub-Saharan Africa is around 49%. He writes that the mean percentage for the R0 allele in the same area is over 65%. Hiernaux regards any marked deviations from these average frequencies as potential indicators of exotic influence in a given population. To his credit, he then attempts to corroborate this with other lines of evidence.
Since Hiernaux already ruled out that the Neolithic pastoralists of East Africa were of Caucasoid stock (unjustifiably, as we saw in part one), preferring instead to assign to these first Afro-Asiatic-speaking settlers a speculative “Elongated African” origin, he is obliged to explain away the many exotic biological influences in the Horn as being recent imports from Southwest Asia. Hiernaux therefore prefaces his serological discussion by remarking that “history and culture show Ethiopia and the Horn of Africa to be strongly impregnated by Arabic influences”. Specifically, he alludes to four waves of settlement from Arabia: one represented by the Sabaeans, whom he suggests established the Axumite Kingdom in the Ethiopian and Eritrean highlands; another by the ancestors of the Gurage; a third wave consisting of Jewish proselytizers; and a final wave comprising the Muslim patriarchs of the various Somali and Afar clans. From this limited perspective, Hiernaux interprets any convergence or divergence of blood group frequencies from the Arabian mean as being equivalent to greater or lesser Caucasoid ancestry, respectively.
Thus, by Hiernaux’s reckoning, the Afar of Djibouti have considerable Arab influence since they have a relatively high frequency of 62% of the M allele, and the “MN blood groups are also useful for assessing a genetic influence from Arabia[…] the M allele has a high frequency in Arabia, above 65 per cent; in sub-Saharan Africa its frequency varies between 31 and 73 per cent”. He asserts that the Galla, Gurage, Amhara, Falasha and Tigre of Ethiopia and Eritrea, who are also Afro-Asiatic speakers, are in a similar position, for “all these populations have high M frequencies of between 63 and 68 per cent”. On the other hand, despite acknowledging strong anthropometric ties between the northern Somalis and Arabs, Hiernaux argues that the northern Somalis (whom he defines as all major Somali clans except for the Sab) have a comparatively weaker serological connection with Arabia because the M allele frequency is a lower 51% among both the Dir Somalis of northwestern Somalia and the Issa Somalis of Djibouti. This happens to be the standard percentage for ethnic Somalis as a whole (see Mourant’s serological Table 1 below). However, what Hiernaux overlooks or is perhaps unaware of is that moderate frequencies of around 50% for the M allele are actually the norm among many other Afro-Asiatic-speaking populations in the circum-Mediterranean area, including Egyptians. In fact, El Hassan et al. (1968) observed an even lower percentage of 48% among the Beja of Sudan. Arthur Ernest Mourant, who pioneered the use of serology and other genetic systems in anthropology, explains the situation thusly:
The frequency of the M gene is high, mostly well above 60 percent, in nearly every Indian population tested, whereas it is below 60 percent and mostly below 55 percent in the Mediterranean area, apart from Sardinia. Insofar as the scanty data available allow us to judge, there appears to be a sudden rise from low, typically European and Mediterranean M frequencies in Turkey (both in Turks and in Ed-Turks) and Lebanon and in the Armenians, to much higher values in Syria, Arabia, Iraq, and Iran. Egyptians show low, typically Mediterranean frequencies, but higher values occur in Sudan, Ethiopia, and Somalia, where they appear on the map as an extension, unique in Africa, of the high M area of Asia.
In other words, what Hiernaux mistakenly interprets as weaker serological Caucasoid affinities for northern Somalis compared to surrounding Afro-Asiatic-speaking groups are actually simply a different type of Caucasoid affiliation (i.e. more Egyptian than Arabian on this particular blood group trait).
As regards his pure “Elongated Africans”, the Tutsi-Hima, Maasai and Ful, Hiernaux is again inconsistent. He is silent on the M allele frequencies for the Maasai and the Ful. Besides the Nama Hottentots of Southern Africa (whom he admits have some ancient Cushitic admixture), he does, though, remark that “the only other sample to show an M frequency above 60 per cent is a small one of Tutsi from Rwanda and Burundi”. Nonetheless, Hiernaux does not propose this unusually high M percentage as evidence of Ethiopian or Arabian introgression into the Tutsi population. He instead maintains his untenable position that the Tutsi Bantus have virtually no exotic genetic influences.
With respect to the Rhesus (Rh) blood group system, Hiernaux writes that the R0 allele’s “frequency varies between 33 and 95 per cent in sub-Saharan Africa, while it never attains the lowest of these values in North Africa or Arabia, and in Europe it does not rise above a few per cent”. He thus asserts that “the R0 frequency is therefore useful for tracing an exotic influence”. On this basis, Hiernaux indicates that “R0 frequency has relatively low values also in northern and central Ethiopia, which has a long history of contacts with southern Arabia”. He notes that these R0 allele percentages are typically near or below 50% among said Afro-Asiatic-speaking populations; at around 48% for the Tigre and Falasha, 54% for the Galla, and 35% for the Gurage. Since the Afar of Djibouti have a similar R0 frequency of 51% whereas the Issa Somalis of the same territory have a much higher R0 frequency of 65%, Hiernaux argues again that Arabian influence is significantly greater among the Afar and the central and northern Afro-Asiatic-speaking Ethiopian groups than among the northern Somalis. But is this really justified?
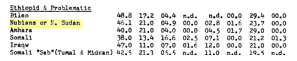
Blood group frequencies of various Afro-Asiatic-speaking populations and Nubians in Northeast Africa. From left: cDe (R0), CDe (R1), cDE (R2), CDue, cDue, Cde, cde (r), cdE (Fleming (1965)).
Hiernaux’s claim is based on R0 frequency data gathered by Fourquet (1970), who analyzed the blood group means of Somalis from a single clan, the Gadaboursi and Issa Dir of Djibouti. However, this percentage appears to have been distorted by genetic drift. This is strongly implied by Iacovacci et al. (2017), who indicate that 100% of their Djiboutian Dir individuals belong to the paternal haplogroup T, while this Y-DNA clade normally occurs at moderate frequencies of ~15% among Somalis from other clans (cf. Sanchez et al. (2005); Immel and Kleiber (2009)). Furthermore, Fleming (1965) reports a much lower R0 frequency of 38% in a general (non-Sab) Somali sample, and percentages of 40% for the Amhara, 48.8% for the Bilen, and 53.8% for the Arsi Galla. Sistonen et al. (1987) likewise observed a ~38% R0 (cDe or DCe) frequency in their large sample of 1026 ethnic Somali individuals in Somalia (cf. Table II). According to Fleming (1979), the blood group data for the Chaha Gurage (which Hiernaux cites) also contains errors in that it mistakes cDE for cDe, thereby reporting lower cDe/R0 frequencies for the Gurage than this population actually has. On balance, going by Hiernaux’s way of thinking, this would suggest that there is greater Arabian serological influence among Somalis than among most other Afro-Asiatic speakers in the Horn. (*N.B. Sistonen et al.’s data is weighted more towards southern Somali individuals, with 59% of their cohort indicating that they were born to southern clans. The scientists, moreover, concede that the southern “Sab are generally considered to be of a more mixed descent, assimilating elements from virtually all of the other clans and, probably, also some from the Galla and Negroid populations the remnants of which still live around the rivers.” This greater Negroid influence explains why, in some of the other serological systems that they examined, their sample set appears more shifted toward hunter-gatherer populations than is usually the case.) Rather than recent Arabian introgression, though, it is more probable that the relatively low cDe/R0 frequencies carried by Somalis were simply bequeathed to them by their Cushitic ancestors (who presumably had an even lower cDe percentage). For one thing, similar cDe frequencies have been observed among some other Afro-Asiatic speakers in North Africa, whom we know from ancient DNA analysis are related to the early Cushites (e.g. Metri et al. (2012) report cDe/R0 frequencies of 31% and 37%, respectively, among Maghrebi individuals from Sid-El-Djilali and M’sirda in the Tlemcen region of western Algeria; Merghoub et al. (1997) likewise reveal a cDe or R0 (Dce) frequency of 32.9% among Mozabite Berbers from Mzab, Algeria; similarly, Skaik (2011) indicates a cDe frequency of 30.4% among Palestinian females in Gaza).

Blood group frequencies among populations in Northeast Africa, Southern Arabia and Southern Europe (Mourant et al. (1976)).
In his landmark thesis The Blood Groups of Somali Tribes published in 1959, K. L. G. Goldsmith collected serological data from over 1000 ethnic Somali individuals inhabiting the former British Somaliland protectorate. His large sample set was drawn from various clans, and is thus representative of the blood group characteristics of northern Somalis in general. Goldsmith observed that the Somalis had an average R0/cDe frequency of 48%, well within the typical range for this allele among the other Afro-Asiatic-speaking populations in the Horn. Mourant et al. (1976) highlight this in their Table 1 (shown to the right).
In his defense, Hiernaux may not have been aware of Goldsmith’s study, as he does not include it among his bibliographical references in The People of Africa. He does, though, cite Goldsmith’s earlier paper from 1958, A Preliminary Investigation of the Blood Groups of the Sab Bondsmen of Northern Somaliland, which I. M. Lewis co-authored. This is quite interesting since Goldsmith and Lewis provide an average R0/cDe frequency for the northern Midgaan and Tumaal low-caste groups, who, along with the Yibir, are collectively known as the Gabooye (not to be confused with the southern Sab or Rahanweyn confederacy). At around 43%, it is almost identical to Fleming (1965)’s R0 value of 42.5% for the same Sab groups. Both are also within the typical allele percentage range of the other Afro-Asiatic-speaking groups in the Horn.

A Gabooye Somali woman. Serological analysis of the low-caste groups among the Somali, who are collectively known as the Gabooye, indicates that they are genetically similar to their higher-caste counterparts. This suggests that the Gabooye are of Cushitic origin like other ethnic Somalis, but probably became differentiated on the basis of their non-pastoralist occupations.
Thus, contrary to Hiernaux’s assertion, a value near 40% is the actual mean percentage for the R0 allele among northern Somalis, including the low-caste groups. Elizabeth W. Ikin, who co-wrote a series of influential blood group studies with Mourant on various populations in Ethiopia, further confirms this in her own 1959 study on blood group distribution in the Middle East. She therein remarks that the average Somali R0 frequency is around 43%, apparently in allusion to Goldsmith and Lewis’ analysis. In his 1980 book Human Biological Diversity co-authored with Amitabha Basu, Hiernaux himself would eventually acknowledge the relatively low R0 frequencies found among both ethnic Somalis and the Beja (perhaps he had since gotten wind of Goldsmith, Ikin and Mourant’s serological work).
With regard to the Rhesus frequencies among the Tutsi Bantus and their Hima kinsmen as well as the Maasai Nilotes and Ful West Africans, Hiernaux is both more and less straightforward. He writes that “moderate frequencies of R0 [are] found in some populations in which an exotic influence has been suspected, in the pastoral Ful of northern Nigeria and in the Hima of Ankole (Uganda)”, yet asserts elsewhere that “as a whole, the Ful and the non-Ful of West Africa are very similar in their allele frequencies for blood trait systems”. Hiernaux also notes that the Ful from Senegal, the Tutsi and the Maasai all have high R0/cDe frequencies like the Hutu and most other populations below the Sahara. This is in direct contrast to Excoffier et al. (1987), who, on the basis of a lack of the sickle-cell trait and apparently lower R0 frequencies than other Bantus (percentages which the authors never divulge), suggest that the “Tutsi and Hima [are] surrounded by Bantu populations but closer genetically to Cushites and Ethiosemites, [and] are known to have migrated from northern territories recently”. However, Hiernaux’s assertion is somewhat in agreement with Fleming (1979), who observes that:
In Rhesus and other systems, however, it is clear that none of the major contemporary Kenyan populations can be classified as Ethiopid, except for the Oromo and Somali. Neither Luo, nor Masai, nor Kikuyu emerge as anything but ordinary members of Nilotic and Bantu clumps.
Similarly, Lawrence Oschinsky (1960) observed:
The Batutsi are tall, narrow nosed, narrow faced Negroids showing evidence of previous Caucasoid admixture, the Bahutu are intermediate in stature and have broader noses and faces, and the Batwa represent a local variant of Congo pygmy racial type. In the ABO, Rh and MN blood group distributions the Batutsi and the Bahutu are similar.
J. D. Fage (2013) likewise indicates that:
The origin of the Tutsi/Hima/Chwezi ruling class in the lacustrine Bantu kingdoms is an intriguing question. Serologically they are Blacks, and this seems to rule out the possibility of a Cushitic origin.
Despite emphasizing the importance of serology in understanding the origins of a given population, Hiernaux does not touch on the ABO blood group systems that are distributed in Northeast Africa. This is regrettable since the O allele that is found at especially high frequencies in Arabia is almost as common among Somalis, Beja and certain other Horn communities as well as among the Berbers of the Maghreb. Coupled with a low occurrence of the more typical African markers (such as the P1 blood group; see Mourant’s Tables 1 and 2 above), this altogether points to a strong serological connection between the Afro-Asiatic-speaking populations on either side of the Red Sea. Mourant (1983) writes:
The main feature of ABO distribution in the region as a whole is the marked westward diminution in the frequency of O from the high levels found on the east coast together with a rise of B and, in certain areas, especially Ethiopia, a rise of A. The high O and low B, especially in the Beja of the eastern Sudan and the Somalis, presumably indicate Arabian admixture in the population concerned, as might be expected from what we know of their history.
El Hassan et al. (1968), on which Mourant served as a co-author, similarly notes that:
In two respects the Beja differ from most of the surrounding peoples, namely in their low frequencies of B and of M. In their low frequency of B they resemble the southern Arabs and the Somalis, though they differ from both in their much higher frequency of all the typical African marker genes. In their lower frequency of M they resemble, on the other hand, some of the African peoples of north-east Africa who possess few of the typical Mediterranean genes, but also the Somalis, who possess many such genes.
Ikin (1959) goes further and asserts that the serological ties between Somalis and Southern Arabs are such that they form their own “Arabian” sub-class of the “Mediterranean race”:
The peoples of the Near East thus fall into two main classes. We have the Turks and Eti-Turks, whose relations appear to be mainly with Europe, and especially with the Mediterranean area. They have a high A gene frequency, a moderately high M gene frequency, together with a high R1, and a fairly high rh-negative frequency. Then we have the “Arabians”, a sub-class of the “Mediterranean” race. These are the Yemenite Arabs and Jews, the Zabidi Arabs and the Socotrans and, with reservations, the Somalis. These all have low A and high O gene frequencies, mostly high R1 and rather lower rh-negative gene frequencies. They also have a very high M frequency which is possibly distinct from the general high M of Asia. It is interesting to note that the Berbers, the “white” race of N. Africa, resemble the Arabs in having a high O frequency, but differ from them in having high N and high Rh-negative frequencies.
Froment (1994) likewise confirms that the blood group traits of Somalis, Afars and other Afro-Asiatic-speaking populations of the Horn, as cataloged in Mourant’s 1976 tome The Distribution of the Human Blood Groups and Other Polymorphisms, show considerable Arabian affinities.

Phylogenetic tree of the blood group affinities of the Afro-Asiatic, Khoisan, Niger-Congo and Nilo-Saharan-speaking populations in Africa. The Afro-Asiatic speakers form two clusters: the first joins Libyans, Berbers and Bedouins of North Africa with Somalis of the Horn at a bootstrap value of 75.6%; the second comprises Tigre, Amhara and Afar of the Horn, with Tigre and Amhara joined at a bootstrap value of 50.8% (López (2013)).
In 2013, Janire Allende López of the Universidad del País Vasco compared the blood group traits of a large selection of Afro-Asiatic, Khoisan, Niger-Congo and Nilo-Saharan-speaking communities inhabiting Africa. Like Mourant, El Hassan, Ikin and Froment before him, López observed that the populations generally grouped along linguistic lines. The Afro-Asiatic speakers formed two such phylogenetic clusters: the first grouping comprised the Libyans, Berbers and Bedouins of North Africa and the Somalis of the Horn of Africa, joined together at a bootstrap value of 75.6%; the second grouping consisted of the Tigre, Amhara and Afar of the Horn, with the Tigre and Amhara sharing a branch at a bootstrap value of 50.8%. The Nilo-Saharan-speaking Baria/Nara of Eritrea also fell into the latter Afro-Asiatic cluster, evidently due to their documented intermixture with local Cushitic- and Ethiosemitic-speaking peoples (cf. Trombetta et al. (2015), Supplementary Table 7; around 60% of Nara individuals today carry the E1b1b modal Afro-Asiatic paternal haplogroup on account of these contacts). On the other hand, the Maasai Nilotes in López’s sample set clustered with other Nilo-Saharan and Niger-Congo populations, just as Fleming had earlier remarked:
Las poblaciones de afroasiáticos, excepto los Funji, se encuentran agrupadas. Por un lado observamos las poblaciones de Libios y Bereberes unidos en una rama con un valor bootstrap de 51% y a los Libios, Bereberes y Beduinos con un valor bootstrap de 75,6%, todos ellos relacionados a los Somalies. Mientras que los Tigre, Amhara, Afar y Baria/Nara se encuentran en otra rama, las poblaciones Tigre y Amhara están unidas en una rama con un valor bootstrap de 50,8 %. Los pigmeos (Mbuti y Biaka) y los Hadza están relacionados con un valor bootstrap de 73%. En cuanto a la familia Khoi-Sanida, se observa que los Khoi y los San están en la misma rama. Por último las familias de Niger-Congo y Nilo-Saharianos se agrupan en el centro del árbol filogenético.
Afro-Asiatic populations, except the Funji, are grouped. On the one hand we observe the populations of Libyans and Berbers united in a branch with a bootstrap value of 51% and the Libyans, Berbers and Bedouins with a bootstrap value of 75.6%, all related to the Somalis. While the Tigre, Amhara, Afar and Baria / Nara are in another branch, the Tigre and Amhara populations are united in a branch with a bootstrap value of 50.8%. Pygmies (Mbuti and Biaka) and Hadza are related with a bootstrap value of 73%. As for the Khoi-San family, it is noted that the Khoi and the San are on the same branch. Finally, the families of Niger-Congo and Nilo-Saharan are grouped in the center of the phylogenetic tree.
Ecology, rock art and genetics
The final pillar of Hiernaux’s “Elongated African” theory focuses on ecology. Specifically, he argues that the linear physiques that are common among the populations inhabiting the southern rim of North Africa evolved as long-term morphological adaptations to hot and arid biotopes or habitats. He writes:
In the hot and dry zones of sub-Saharan Africa, selection has favoured a different body build, which also displays a low mass-to-surface ratio : a gracile, elongated physique, with long legs and narrow shoulders. Weight is relatively very low in such people of medium or tall stature, but not especially so in absolute value. This tendency of the human physique toward tallness in the hot and dry zone explains the failure of Bergmann’s rule in sub-Saharan Africa. Like size reduction, it represents an adaptation to heat, but this time in a dry climate in which cooling by sweating is the easiest.
Hiernaux stresses that head, face and nose form is also linked with environmental variables:
Head and face shape also are significantly correlated with air temperature and moisture in sub-Saharan Africa. The more extreme are heat and dryness, the narrower tends to be the head. Face width tends to increase with air moisture and climatic uniformity. Nose height tends to be lower in wetter regions. Of all measurements, nose width shows the closest correlations with climate: it tends to increase with the annual rainfall and to decrease with the temperature of the hottest month.
On the surface, the various anthropometric tables that Hiernaux provides in The People of Africa indeed appear to support his theory on the correlations between body dimensions and habitat. That is, until one closely examines his data table on the “pure” northern Nilotes of the Nile Valley:
 As can be seen in the anthropometric means above, while all of Hiernaux’s Nilotic populations are indeed tall and possess very narrow heads and relatively narrow faces, their nose heights and nose breadths are not what one would expect from individuals whose ancestors supposedly evolved in a hot and dry habitat. Instead, they have wide and flat noses like the average Bantu and West African population — groups that Hiernaux insists evolved in a moist, jungle environment — with the platyrrhine nasal indices to match. This also contradicts Hiernaux’s assertion that “obviously a narrow face could not accommodate a very wide nose” since the Nilotes do, in fact, possess both morphological traits at once. Altogether, the anthropometric means suggest that the Nilotes evolved in a forested biotope similar to that which molded the Bantus, West Africans and Pygmies, with the main difference being that the Nilotes likely inhabited a sunnier portion of the same area.
As can be seen in the anthropometric means above, while all of Hiernaux’s Nilotic populations are indeed tall and possess very narrow heads and relatively narrow faces, their nose heights and nose breadths are not what one would expect from individuals whose ancestors supposedly evolved in a hot and dry habitat. Instead, they have wide and flat noses like the average Bantu and West African population — groups that Hiernaux insists evolved in a moist, jungle environment — with the platyrrhine nasal indices to match. This also contradicts Hiernaux’s assertion that “obviously a narrow face could not accommodate a very wide nose” since the Nilotes do, in fact, possess both morphological traits at once. Altogether, the anthropometric means suggest that the Nilotes evolved in a forested biotope similar to that which molded the Bantus, West Africans and Pygmies, with the main difference being that the Nilotes likely inhabited a sunnier portion of the same area.
On some level, Hiernaux appears to be aware of the common origin of the Nilotic, Bantu and West African populations, for he alludes to a “most numerous and most widely spread African stock which prevails in West and Central Africa”. He indicates that this “West Central African” ancestral stock (essentially a euphemism for the “Negroid” taxon of traditional physical anthropology) is best represented in the archaeological record by populations from the Mesolithic Wadi Halfa in Sudan and the Neolithic and protohistoric southern Sahara, such as the Asselar and Iwo Eleru specimens.
Hiernaux also concedes that the aforediscussed “East African skeletal material is not closely related to the Mesolithic Wadi Halfa remains”, yet speculates that “these people may have evolved from the stock represented by the Wadi Halfa population”. But is there any evidence to support this association? As discussed in detail on Ancient DNA from Sudan, the Mesolithic inhabitants of Wadi Halfa (the immediate ancestors of the Nilotes) are instead both genetically and morphologically distinct from the later A-Group, C-Group and Kerma and Meroitic, Post-Meroitic/X-Group and Christian period inhabitants of the Nile Valley (the ancestors of the Afro-Asiatic-speaking populations in Northeast Africa). Hiernaux is therefore mistaken about that as well.
So where, then, does Hiernaux believe his “Elongated African” morphology originated? He postulates that it developed in the Neolithic Sahara among early pastoralists, and subsequently spread from there during one of the region’s various dry periods. The migrating herders would then have undergone further local evolution in the arid zones of East Africa and the Nile Valley. As mentioned earlier in the essay, Hiernaux suggests that these Saharan pastoralists are represented by the C-Group people. This is quite ironic since, as we also saw, the C-Group herders were of Caucasoid physical type and had non-kinky hair form. They therefore were not the Tutsi- or Maasai-like “Elongated Africans” that Hiernaux had conjectured. On this ancient settlement of Saharan pastoralists, he indicates:
Quite possibly the migration of the Sahara pastoralists reached the Horn and East Africa. Settlements of village farmers, basically neolithic but also using some copper, are known from the plateau of north-western Ethiopia; their material culture shows affinities with that of the C-Group peoples who moved into Nubia from the western desert about 2500 BC. Rock paintings in the eastern parts of Ethiopia and Somalia are reminiscent of those of the eastern Sahara; they depict herdsmen and long-horned, humpless cattle.

Global racial distribution during the Middle Ages (Coon (1982)).
Additionally, in reference to his hypothetical “Elongated African” morphology, Hiernaux writes that “Coon believes that it was first concentrated in the cattle breeders of the neolithic Sahara, whose inhabitants dispersed because of progressive desiccation, giving rise to the herdsmen of East Africa”. However, this passage is rather misleading since Coon never made any such assertion or indeed even acknowledge an “Elongated African” entity. He instead maintained throughout his career that the first Afro-Asiatic-speaking settlers in the Horn region and environs were “Mediterranean Caucasoids”, who were ancestrally related to the “Hamitic” predynastic Egyptians and ancient Libyans. In 1982, Coon published Racial Adaptations, an update of his earlier Races: A Study of the Problems of Race Formation in Man from 1950. The latter was among the first scientific works to explore biological adaptation in human populations. Although Racial Adaptations was released after Hiernaux’s own book on the subject, Coon was not swayed by his colleague’s theories, as the enclosed racial distribution map makes clear.
Ultimately, Coon regarded the Middle East rather than the Sahara or East Africa as the likeliest area of origin of the Mediterranean stock. He explains it thusly in his The Races of Europe:
whatever the date of these specimens in years, East Africa was not, in Upper Palaeolithic times, the center of Mediterranean racial evolution. Neither, it would appear, was the Sahara; so far the archaeologists have not found evidences of the Upper Palaeolithic Capsian culture in the central zone of the desert itself, where there is at present a gap between the Levalloisian and what appears to be an early, arrow-chipping Neolithic in Capsian tradition. The Capsian apparently came to North Africa from the east, and the mid-Sahara may have served even during Pleistocene times as a dividing line between white and negroid humanity, just as it does today.
-
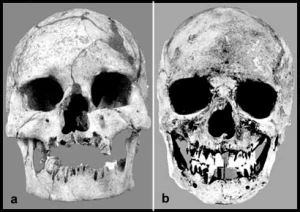
-
Iberomaurusian (left) and Capsian (right) skulls excavated in North Africa.
-
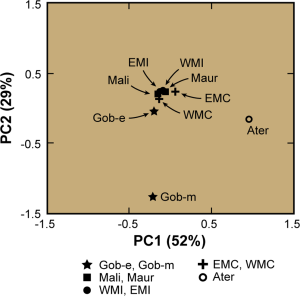
-
Craniometric analysis of Iberomaurusians (WMI, EMI), Capsians (EMC, WMC), “Mechtoids” from Mali and Mauritania (Mali, Maur), early Holocene Kiffians from Gobero (Gob-e), mid-Holocene Tenerians from Gobero (Gob-m), and ancient Aterians (Ater). The Iberomaurusian, Capsian, Mechtoid and Kiffian samples cluster together, showing Upper Paleolithic European ties. The Tenerian cohort sits apart, likely because of its “Mediterranean” affinities. The Aterian specimens, which are the oldest at over 40,000 years of age, are also an outlier; this is apparently due to their proposed Khoisan-related affiliation (Sereno et al. (2008)).
(For further details, see Did the Capsians of North Africa descend from the Iberomaurusians?, Have the Kiffians and Tenerians of the Gobero been genetically analysed yet?, and What are the genetic affinities of the Aterians?.)
Besides the foregoing, there are several additional key indications that Coon, as opposed to Hiernaux, is factually closer to the mark about the identity and provenance of the first Afro-Asiatic-speaking herders in the Horn region.
Firstly, ecological studies have established that the climate and lake levels in the Horn fluctuated during the Holocene (~10,000-6,000 ybp). The highlands were initially frigid and uninhabitable. This early wet phase was characterized by a glacial retreat, with a corresponding rise in the water levels of the Ethiopian and East African Rift Valley lakes as the snow melted. These lake levels would remain high throughout the Holocene, thereby precluding human occupation. As the climate gradually changed and the lake levels dropped, vegetation favoring pastoralism would begin to grow. Only after 5,000 ypb, with the start of a new dry phase, would the first Neolithic pastoralists arrive from the Sahara. Oba (2013) writes:
In the region of the Horn of Africa, both the highlands and the lowlands experienced climatic fluctuations. It is possible that during the early phase of the wet Holocene climate, the highlands were too cold for human settlement. Finnevan reports: “The …wet phase was accompanied by a glacial retreat” from the highlands in both Ethiopia and East Africa. Between 12,000 and 5,000 years ago snow retreat resulted in a rise in the levels of the Ethiopian and the East African Rift Valley lakes. At the regional level, the fluctuation in the East African Rift Valley lakes was synchronized, suggesting close links between changes in lake levels and regional climate..[…]
In East Africa, during the full Holocene period from around 10000 through 6000 BP, the high water levels in the Rift Valley lakes did not allow human settlement in the moist environments. The wet and the dry phases alternated between 4450 BP and 2700 BP, with lake levels dropping, followed by recession of the sub-humid vegetation towards arid conditions in the lowlands and the retreat of forest cover in higher elevations. From the Afar region of Ethiopia, lakebed changes depict the different phases of the paleoclimate. Grove, referring to carbon dating of the sediment cores of the lakes (i.e. Shala, Abiyata and Langano), confirms higher lake levels around 9200 BP, which retreated to the present levels by 4000 BP. The decline in the lake levels is an indication that climate conditions had shifted to a dry phase ca. 5000 BP[…]
The Neolithic herders arrived from the Sahara. The period between 5000 BP and 3000 BP represents the mid-Holocene, when aridity induced migrations of prehistoric pastoralists into the Horn of Africa.
The Horn was thus considerably less arid in the past than it is now. This perhaps should be obvious given the presence of certain fauna on the region’s ancient cave paintings, which can only survive in habitats where there is abundant vegetation (e.g. the ibex). Indeed, the very type of cattle depicted in the earliest pastoral rock art, the long-horned humpless cattle or Bos taurus, itself serves as a reliable indicator of the kind of climate that prevailed at the time. Oba explains:
During the Neolithic period called Bovidian, the styles of the rock art enable various interpretations based on the depiction of livestock species such as ovids (goats) and capris (sheep) and Bovin (cattle). By about 8000 BP, the depictions in the rock art were mainly of ovicaprids and cattle. The rock artists were predominantly concerned with the sources of their food during each period. Thus, the shift from the Paleolithic phase, which represented the hunter foragers, to Neolithic developments, which represented pastoralists, show shifts in food sources and society responses to climate and environmental changes. The domestication of Bos primigenius (primitive cow) expanded into the Nile valley about 7000 BP; this was the ancestor of the cattle breeds Bos taurus (the humpless long horned) and Bos indicus (the humped short horned zebu). Bos taurus was common during the humid phase of the climate, while Bos indicus became prominent with increased aridity. The depictions show that the rock art serves as a slow cinematic sequence of pastoral evolution to climate change, even when data from a single site is scrutinized. In particular, the gradual disappearance of Bos taurus and the popularity of Bos indicus in rock art sites strongly imply environmental adaptation since Bos indicus had better thirst tolerance than its predecessor. The camel was introduced later than the bovids during periods of increased aridity. From the rock art across the region archaeologists have pieced together the regional migration patterns of the rock art pastoralists.
Recent archaeological excavations in northern Somalia, during which the first actual artifacts from the Land of Punt were unearthed (as explained in detail here), provide a similar indication. Among the discovered objects was an anthropomorphic crocodile figurine of apparent religious significance. This suggests that the ancient Puntites inhabited a more lush environment than today because crocodiles usually congregate in freshwater biotopes.
Hiernaux was therefore mistaken when he proposed that the Horn was once dry enough for the “Caucasoid” morphology to have, at least in part, evolved in situ there. Since the region would not become arid until relatively recently, the ancient pastoralists with that phenotype had clearly arrived from elsewhere.
Secondly, the pastoral-themed cave paintings at Laas Geel, Dhambalin and Karin Hegane in northern Somalia and at other sites in Northeast Africa are in a singular “Ethiopian-Arabian” style. The earliest examples of this rock art tradition are found in Saudi Arabia, and they date back to around 4,500 BCE. On this point, Rodolfo Fattovich of the Università l’Orientale di Napoli, who in 2001 led archaeological excavations at Mersa/Wadi Gawasis on the Red Sea (the ancient Egyptian port of Saww used for commercial expeditions to the Land of Punt), notes:
Finally, a strong interregional interaction, if not a proper movement of people from Arabia to Ethiopia, in the 3rd-2nd millennia B.C. is supported by the occurrence of rock pictures in a typical style in both regions. This style is characterized by painted or engraved bovines with the body in profile and the head and horns in plan. It is recorded as «Jubba Style» in northern Saudi Arabia, «Dahthamani Style» in central Arabia, and «Karora Style» or «Ethiopian-Arabian Style» in the Horn of Africa.
The earliest evidence of this style has been traced in northern Saudi Arabia and dated to the mid-5th millennium B.C. By the mid-3rd millennium B.C., most likely, it spread to the southern Hidjaz (Saudi Arabia), northern Hararge (eastern Ethiopia), and – along the Ethiopian Rift Valley – to Sidamo (southern Ethiopia). Then, figures dating back to the 2nd-early lst millennia B.C. have been recorded at Jebel Qara (central Arabia), Hararge (eastern Ethiopia), northern Somalia, and Eritrea. By the lst millennium B.C. this style spread from Eritrea to Nubia, southern Upper Egypt and the Sahara.
Thirdly, the pre-Islamic religion of the Cushitic Somalis is related to that adhered to in the pre-Islamic Middle East. Mukhtar (2003) notes that the Somali word for spirit, hoobal, is derived “from the Arabic hubal, a pre-Islamic Arabian god whose effigy was once venerated in the Ka’ba in Mecca.” Ali (2006) similarly indicates:
the Biblical YAHWE (later turned Yehova and Jehova) was evidently the same as the Somali YAHU – traditionally invoked to ward off evil or danger. While the Cananite god ‘Pal’ was still present in Somalia in the same sense in one or two words, the ancient Aramaic name for the almighty, EBBE, was to this day the most commonly used names for God besides the Islamic ‘Allah’. The Biblical TUBAN-CAIN, whose profession was to make instruments (Genesis 4:22) was obviously a Greek mispronunciation of TUMAL, the Somali iron-monger.
Fourthly, Tishkoff et al. (2010) reports that the most common lactase persistence allele carried by East Cushitic individuals is G-13915. This variant peaks among Somalis at a frequency of 50%. It is closely associated with camel pastoralism and is believed to have evolved in the Arabian peninsula, where the mutation is today prevalent. To this end, Fattovich (1997) asserts that “an introduction of camel from Arabia into the Horn at an early time, perhaps between 2500 and 1500 B.C., might be also supported by the use of an archaic South Arabian type of camel saddle in Somalia and Socotra.”

Neolithic cave paintings at Laas Geel depicting long-horned humpless cattle (the “Hamitic Longhorn”).
Fifthly, analysis by Decker et al. (2014) of 134 cattle breeds from around the world has established that the long-horned humpless cattle depicted in the Neolithic cave paintings of Northeast Africa — cattle that is often colloquially referred to as the “Hamitic Longhorn” because of its ubiquity in Ancient Egypt — is, in fact, related to cattle that was first domesticated in the Middle East at an earlier date (~10,000 ypb). The cattle was therefore brought to Africa by migrating herders, who not long afterwards began drawing the “Ethiopian-Arabian” style rock art in the areas in which they settled:
Geneticist and anthropologists previously suspected that ancient Africans domesticated cattle native to the African continent nearly 10,000 years ago. Now, a team of University of Missouri researchers has completed the genetic history of 134 cattle breeds from around the world. In the process of completing this history, they found that ancient domesticated African cattle originated in the “Fertile Crescent,” a region that covered modern day Iraq, Jordan, Syria and Israel. Lead researcher Jared Decker, an assistant professor of animal science in the MU College of Agriculture, Food and Natural Resources, says the genetics of these African cattle breeds are similar to those of cattle first domesticated in the Middle East nearly 10,000 years ago, proving that those cattle were brought to Africa as farmers migrated south. Those cattle then interbred with wild cattle, or aurochs, which were native to the region, and changed their genetic makeup enough to confuse geneticists[…]
“In many ways, the history of cattle genetics mirrors human history,” Decker said. “In the case of African cattle, anthropologists and geneticists used to suspect that domesticated African cattle were native to the continent, when in fact, they were brought by migrating peoples thousands of years ago. By better understanding the history of the animals we domesticate, we can better understand ourselves.”
Mwai et al. (2015) provide a useful map showing how the cattle likely spread after their initial introduction from the Fertile Crescent:
 Lastly, genetic analysis of modern and ancient human populations in Northeast and Northwest Africa also supports Middle Eastern affinities for the continent’s first Afro-Asiatic-speaking settlers. Hodgson et al. (2014) found a West Eurasian ancestral component that defines the Afro-Asiatic speakers in the Horn, with a frequency peak among ethnic Somalis. Dobon et al. (2015) observed an analogous element among Egyptian Copts, Beja and other Afro-Asiatic speakers in the Nile Valley and Ethiopia, as well as among many present-day Nubians. Henn et al. (2012) in turn identified a West Eurasian ancestral component that defines the Afro-Asiatic-speaking populations in the Maghreb, with a frequency peak among Tunisian Berbers. In addition, as explained in detail in the Ancient DNA from Sudan thread, Sirak et al. (2015) noted a similar Middle Eastern affiliation for the ancient DNA of a specimen from the medieval site of Kulubnarti near the Nile. (*N.B. In 2023, researchers from Uppsala University analysed the DNA of Middle Neolithic pastoralists buried at Skhirat, an archaeological site in northern Morocco. The scientists report that the Skhirat individuals bear Levantine ancestry (cf. Simões (2023a); Simões (2023b)). Our genome analysis of modern Cushitic, Ethiosemitic and North Omotic speakers from the Horn of Africa has found that these individuals also carry varying degrees of the Skhirat Levantine ancestry, with a percentage climax among the Cushitic-speaking pastoralist groups. The Cushites of the Pastoral Neolithic and the medieval Nubians likewise harbor this same Levantine ancestry. Hence, the Middle Neolithic specimens from Skhirat, who originated from the Middle East, appear to belong to the Afro-Asiatic-speaking population that first introduced pastoralism to Africa. For further details, see Ancient DNA from Sudan.)
Lastly, genetic analysis of modern and ancient human populations in Northeast and Northwest Africa also supports Middle Eastern affinities for the continent’s first Afro-Asiatic-speaking settlers. Hodgson et al. (2014) found a West Eurasian ancestral component that defines the Afro-Asiatic speakers in the Horn, with a frequency peak among ethnic Somalis. Dobon et al. (2015) observed an analogous element among Egyptian Copts, Beja and other Afro-Asiatic speakers in the Nile Valley and Ethiopia, as well as among many present-day Nubians. Henn et al. (2012) in turn identified a West Eurasian ancestral component that defines the Afro-Asiatic-speaking populations in the Maghreb, with a frequency peak among Tunisian Berbers. In addition, as explained in detail in the Ancient DNA from Sudan thread, Sirak et al. (2015) noted a similar Middle Eastern affiliation for the ancient DNA of a specimen from the medieval site of Kulubnarti near the Nile. (*N.B. In 2023, researchers from Uppsala University analysed the DNA of Middle Neolithic pastoralists buried at Skhirat, an archaeological site in northern Morocco. The scientists report that the Skhirat individuals bear Levantine ancestry (cf. Simões (2023a); Simões (2023b)). Our genome analysis of modern Cushitic, Ethiosemitic and North Omotic speakers from the Horn of Africa has found that these individuals also carry varying degrees of the Skhirat Levantine ancestry, with a percentage climax among the Cushitic-speaking pastoralist groups. The Cushites of the Pastoral Neolithic and the medieval Nubians likewise harbor this same Levantine ancestry. Hence, the Middle Neolithic specimens from Skhirat, who originated from the Middle East, appear to belong to the Afro-Asiatic-speaking population that first introduced pastoralism to Africa. For further details, see Ancient DNA from Sudan.)
In conclusion, Hiernaux was initially on the right path when he acknowledged an Afro-Asiatic influence of varying degrees on other populations in Africa. Where he went wrong was in allowing unfortunate post-colonial circumstances, both in his place of residence in eastern Central Africa and in academe, to force him to sacrifice empirical truth for political correctness and conjecture. As techniques on how to successfully extract ancient DNA continue to be refined and more old human remains are analyzed, other speculative fallacies will similarly fall by the wayside while robust traditions will instead receive confirmation.





















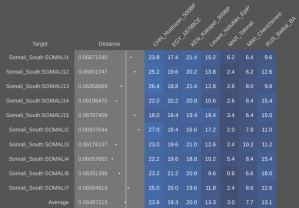
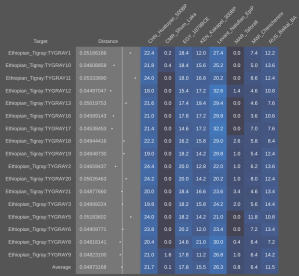







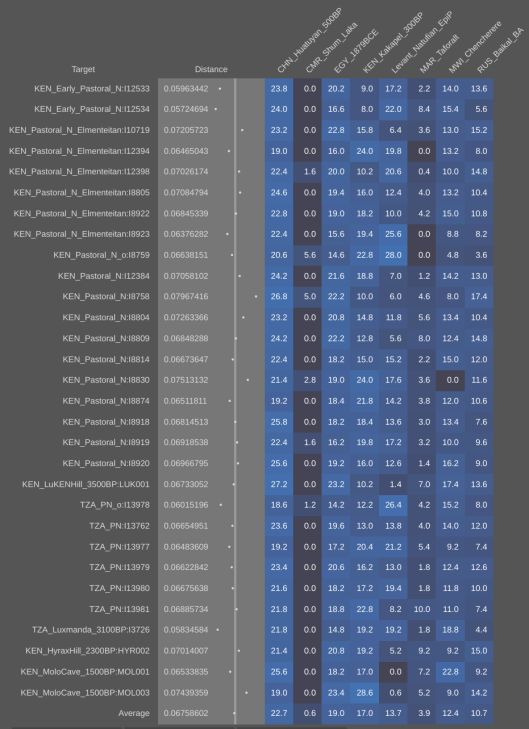

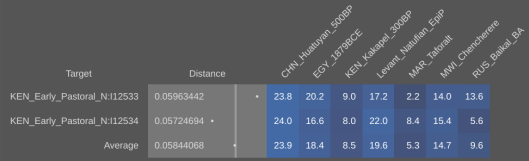




























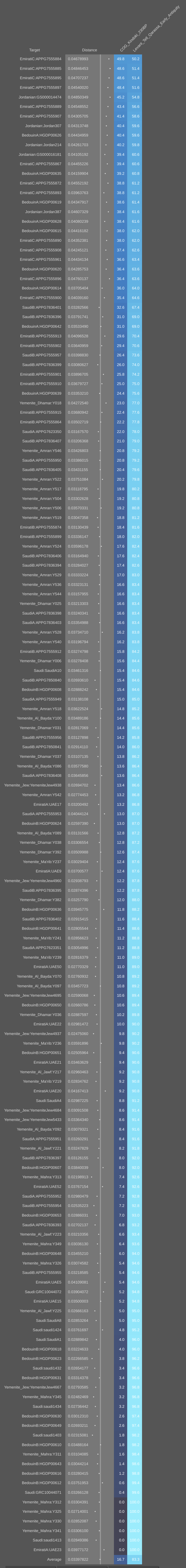
















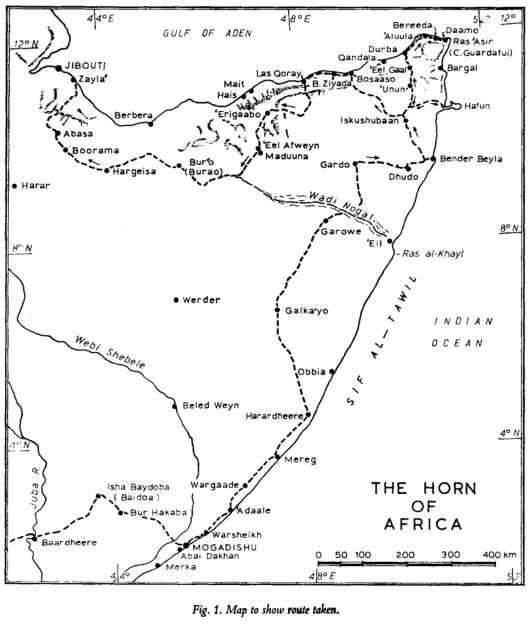








![Closeup of the original single panel in the Tomb of Seti I. The mural depicts (from left) “black” Nubian, copper-brown Egyptian, yellow-brown Levantine, and white Temehu figures. (*N.B. This is supported by archaeogenetic analysis. Haber et al. (2017) note that Bronze Age Levantine individuals from Sidon generally looked like their modern descendants, but had a darker complexion: "SNPs associated with phenotypic traits show that Sidon_BA and the Lebanese had comparable skin, hair, and eye colors[...] but with Sidon_BA probably having darker skin than Lebanese today from variants in SLC45A2 resulting in darker pigmentation".)](https://landofpunt.files.wordpress.com/2016/02/tomb-of-seti-i-single-panel-original-mural.jpg?w=300)










































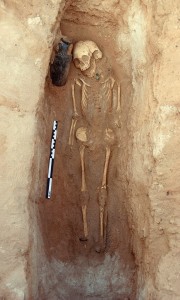




















![A Somali man of "pure" Cushitic type, also showing little Sub-Saharan African influence. Notice the soft-textured close-cropped hair. Clark indicates that "during [Amenhotep II's] reign and earlier, in that of Tuthmosis III, an intermediate 'bobbed' hairstyle appears, and thereafter Puntites have close-cropped hair similar to that of the chief of Punt under Hatshepsut."](https://landofpunt.files.wordpress.com/2023/08/somali-man-cushitic-194a.jpg?w=529)


















































































































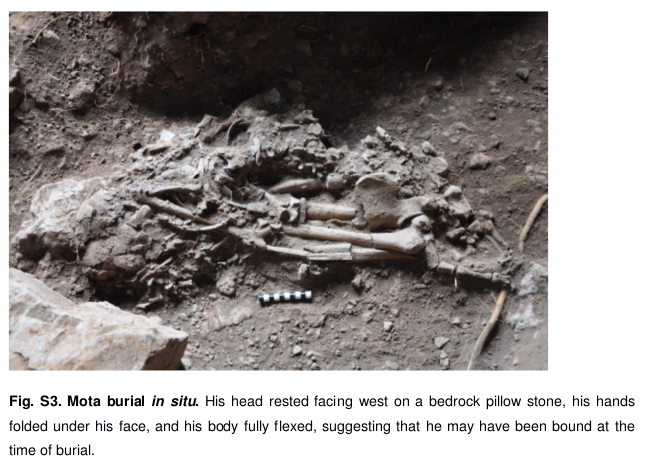
























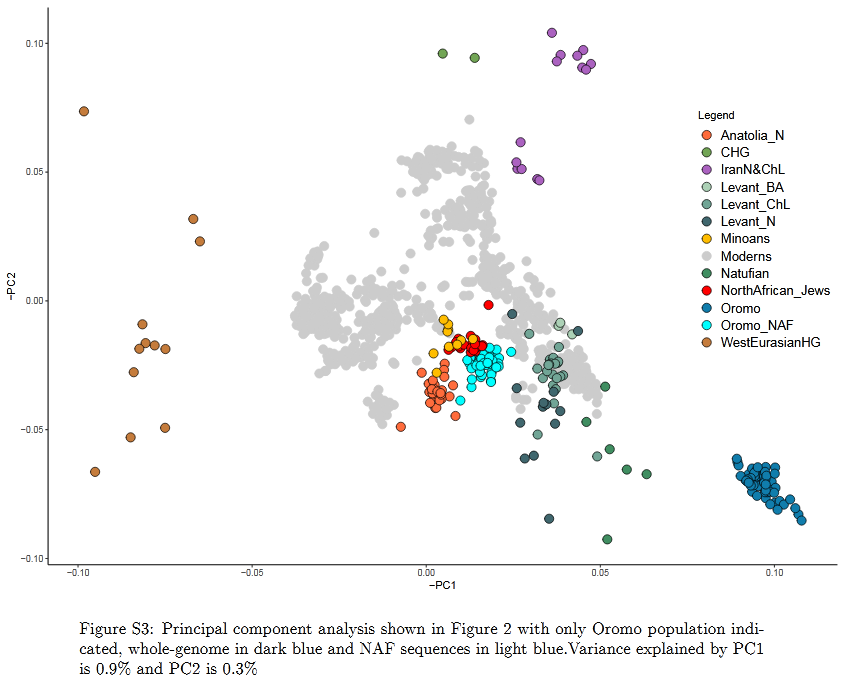













































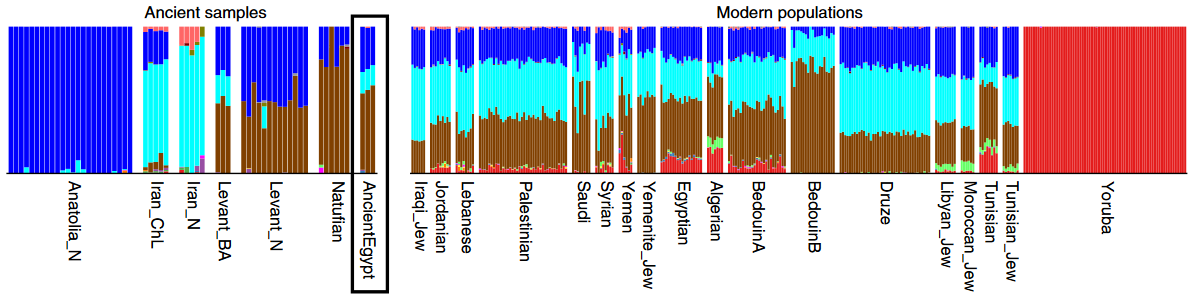


































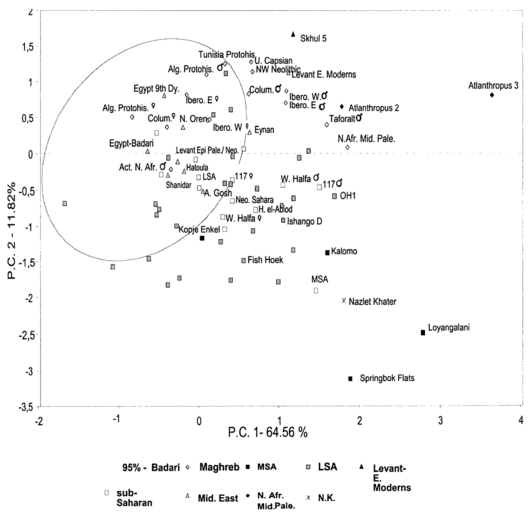
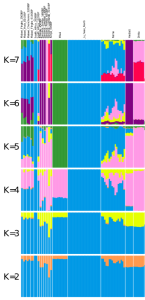
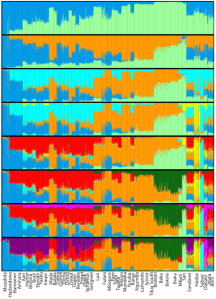
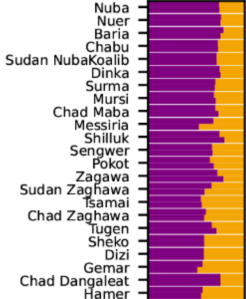


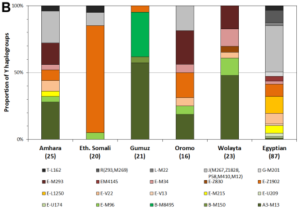
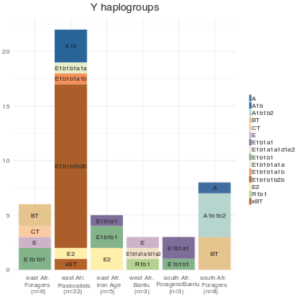




























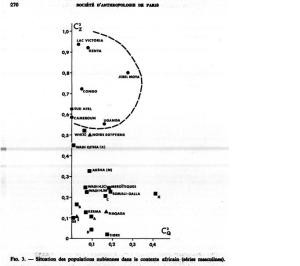

































































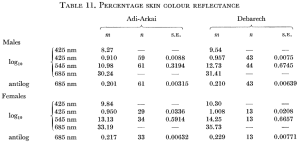
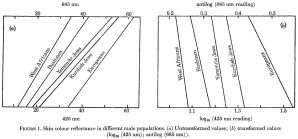















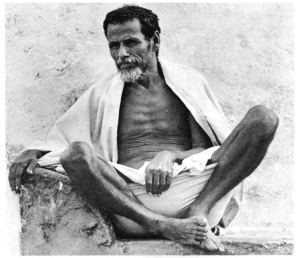














![Berber dancer and actress Sofia Boutella. Note the close likeness to Jawahir Ahmed. This calls to mind Coon (1939)'s assertion that "the Tuareg in their pure form belong to a specialized Mediterranean sub-type[...] They resemble the East African Hamites very closely, and especially the whiter element among the Somali."](https://landofpunt.files.wordpress.com/2015/11/berber-beidan.jpg?w=209)












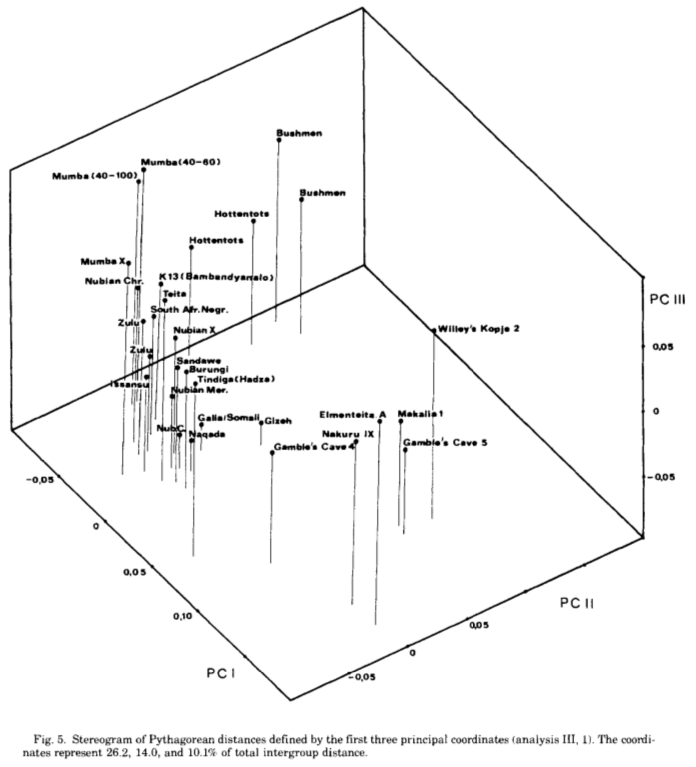




























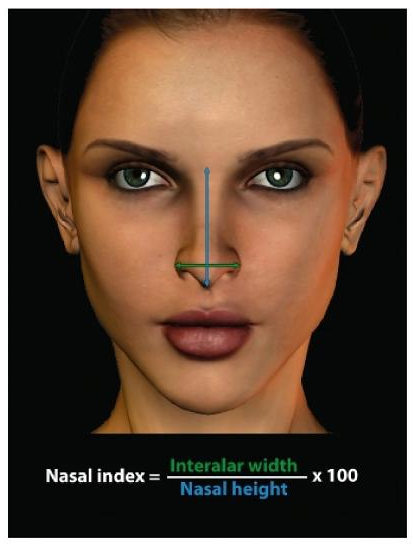


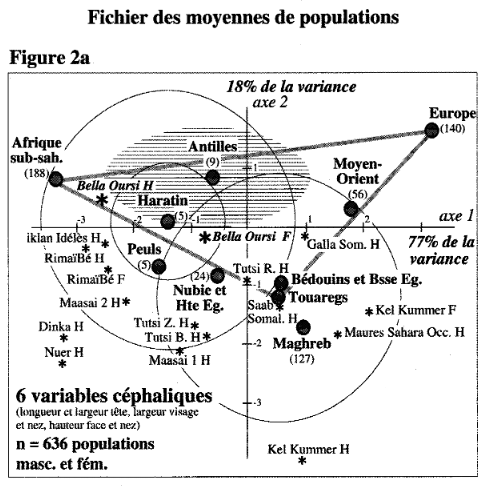

































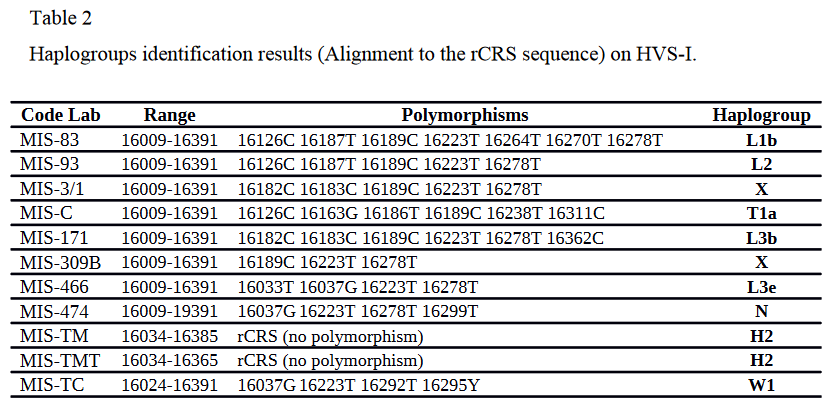











































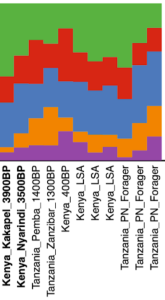
![Genetic modeling of medieval Swahili individuals. Most of the examined individuals derive about 60% of their ancestry from the earlier Bantu individuals from Makwasinyi, with the rest of their ancestry primarily derived from Persian sources and secondarily from Indian sources. Brielle et al. (2022) further note that "Makwasinyi associated [ancestry] itself is ~80% Bantu-related and ~20% pastoralist-related," where pastoralist includes ancestry related to the ancient Cushites of the Pastoral Neolithic. Ergo, before the admixture events between Persian settlers and Bantus that established the medieval Swahili culture, the Bantus would have moved from the continental interior, where they were originally based, to the seaboard. There, in their new place of settlement, the Bantus would have encountered and eventually absorbed the Cushitic Azanians, who, as we already know from archaeogenetic analysis of Iron Age skeletons, preceded them on the Swahili coast (Brielle et al. (2022)).](https://landofpunt.files.wordpress.com/2023/03/brielle2022-3.png?w=529)



































































































































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
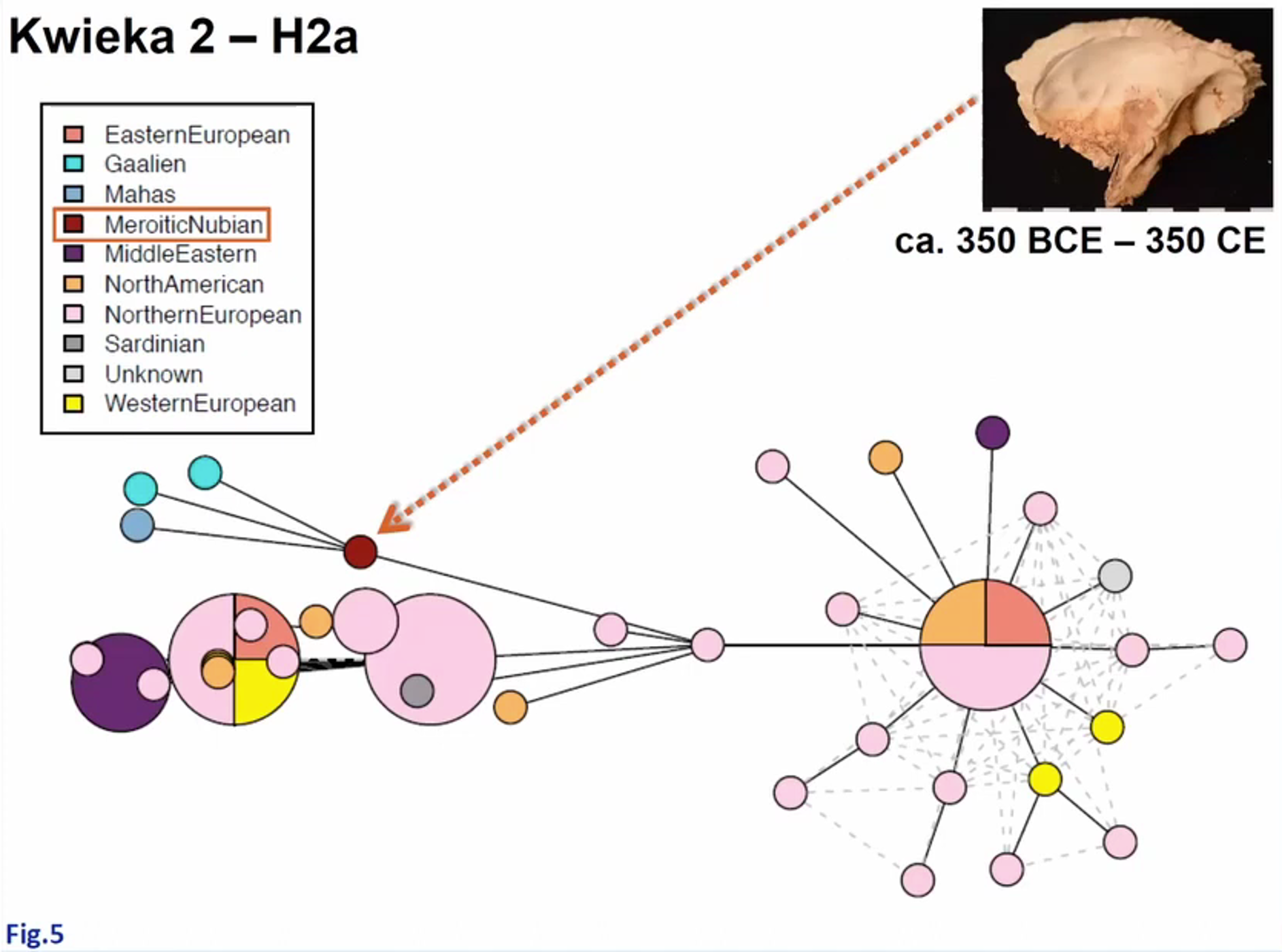{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}